Current issue
Archive
Manuscripts accepted
About the Journal
Editorial office
Editorial board
Abstracting and indexing
Subscription
Contact
Ethical standards and procedures
Most read articles
Instructions for authors
Article Processing Charge (APC)
Regulations of paying article processing charge (APC)
IMMUNOLOGY / STATE OF THE ART PAPER
Ankyrins in human health and disease – an update of recent experimental findings
1
Laboratory of Gastroimmunology, Department of Immunology and Infectious Biology, Faculty of Biology and Environmental Protection, University of Lodz, Poland
2
Department of Cardiology, Bieganski Regional Speciality Hospital, Medical University of Lodz, Poland
Submission date: 2017-10-12
Final revision date: 2018-02-09
Acceptance date: 2018-02-25
Online publication date: 2019-11-18
Publication date: 2020-05-26
Arch Med Sci 2020;16(4):715-726
KEYWORDS
TOPICS
ABSTRACT
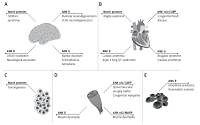
Ankyrins are adaptor molecules that in eukaryotic cells form complexes with ion channel proteins, cell adhesion and signalling molecules and components of the cytoskeleton. They play a pivotal role as scaffolding proteins, in the structural anchoring to the muscle membrane, in muscle development, neurogenesis and synapse formation. Dysfunction of ankyrins is implicated in numerous diseases such as hereditary spherocytosis, neurodegeneration of Purkinje cells, cardiac arrhythmia, Brugada syndrome, bipolar disorders and schizophrenia, congenital myopathies and congenital heart disease as well as cancers. Detecting either down- or over-expression of ankyrins and ergo their use as biomarkers can provide a new paradigm in the diagnosis of these diseases. This paper provides an outline of knowledge about the structure of ankyrins, and by making use of recent experimental research studies critically discusses their role in several health disorders. Moreover, therapeutic options utilizing engineered ankyrins, designed ankyrin repeat proteins (DARPins), are discussed.
| eISSN: | 1896-9151 |
| ISSN: | 1734-1922 |
We process personal data collected when visiting the website. The function of obtaining information about users and their behavior is carried out by voluntarily entered information in forms and saving cookies in end devices. Data, including cookies, are used to provide services, improve the user experience and to analyze the traffic in accordance with the Privacy policy. Data are also collected and processed by Google Analytics tool (more).
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.