
INTRODUCTION
Diabetes is a multisystem disease affecting multiple organs and tissues [1]. While the most recognized microvascular complications involve the kidneys and retina, diabetes is also characterized by peripheral neuropathy and impaired wound healing [2, 3]. Importantly, long-term diabetes also induces microvascular and structural changes in the skin, particularly epidermis and subcutaneous tissue [1, 4]. Although these alterations are often subclinical, approximately one-third of patients develop clinically apparent cutaneous manifestations, including diabetic dermopathy, pruritus, and necrobiosis lipoidica [5, 6].
Chronic hyperglycemia directly affects skin cells and also promotes the formation of advanced glycation end products (AGEs), which contribute to oxidative stress and cellular damage [7]. AGEs exert their effects primarily through binding to the receptor for advanced glycation end products (RAGE), a member of the immunoglobulin superfamily involved in innate immune responses. Upon activation, RAGE triggers intracellular signaling pathways that amplify inflammation and contribute to the pathogenesis of diabetic complications [8, 9].
Aside from AGEs, RAGE interacts with multiple ligands associated with inflammation and oxidative stress, including advanced oxidation protein products (AOPPs), S100 proteins, high-mobility group box-1 (HMGB1), diaphanous-related formin 1 (Diaph1), and lysophosphatidic acid (LPA) [10, 11]. Our previous and ongoing studies have demonstrated the involvement of in diabetic neurovascular complications, such as neuropathy and retinopathy [12–16]; however, its role in diabetic skin disease remains insufficiently explored.
Among RAGE ligands, S100 calcium-binding protein B (S100B) is a key extracellular mediator of proinflammatory signaling. Under physiological conditions, its expression is primarily limited to the nervous system, although it is also found in epidermal melanocytes and resident macrophages i.e. Langerhans cells [17]. Its role in skin pathologies in diabetes remains unclear. Similarly, HMGB1 is a nuclear protein that can be released extracellularly in response to cellular stress, injury, or hyperglycemia, thereby contributing to innate immune activation [18, 19]. Its distribution in the skin appears to change under pathological conditions, suggesting a role in wound healing and inflammation.
OBJECTIVE
In the present study, we focused on the expression of RAGE and its key proinflammatory ligands in epidermis of type 2 diabetic patients to provide a basis for further studies on RAGE-mediated mechanisms in diabetic microangiopathy.
MATERIAL AND METHODS
Patients
Five age-matched healthy volunteers and four male patients with type 2 diabetes were selected for the study. Skin biopsies were obtained from clinically intact skin of the lower leg. Individuals with known major comorbidities or active skin lesions at the biopsy site were not included. All participants provided written informed consent prior to enrollment.
The study protocol was approved by the Institutional Bioethics Committee (No. 10/2010 dated 25 March 2010) and was conducted in accordance with the Declaration of Helsinki (as revised in Fortaleza, Brazil, October 2013). Diabetic patients were diagnosed according to the World Health Organization criteria and were treated on an outpatient basis at a local diabetes center, and received standard diabetes care in accordance with applicable recommendations of the European Association for the Study of Diabetes (EASD) and Diabetes Poland (Polskie Towarzystwo Diabetologiczne, PTD).
Methods
Skin biopsy specimens were obtained under local anesthesia using sterile disposable 3-mm punch instruments from the medial aspect of the lower leg (10 cm proximal to the medial malleolus) on the non-dominant limb [20].
Samples were post-fixed in 4% buffered paraformaldehyde for 10 min, rinsed in phosphate buffer, transferred to 20% sucrose solution for cryoprotection, embedded in Tissue-Tek® O.C.T. Compound (Sakura Finetek, USA), and cryosectioned at –24°C into 10 µm slices. The obtained sections were then processed for immunohistochemical staining using a standardized protocol (Vectastain ABC-HRP kit, Vector Laboratories, USA).
The following primary antibodies were used: rabbit anti-RAGE (Abcam, ab216329, 1 : 200), rabbit anti-CML (Carboxymethyllysine, advanced glycation end-product; Abcam, ab125145, 1 : 200), rabbit anti-S100B (Abcam, ab52642, 1 : 200) and rabbit anti-HMGB1 (Abcam, ab18256, 1 : 200). Negative controls were performed using omission of the primary antibody.
Stained sections were visualized using light and fluorescence microscopy (Olympus-Evident, Japan) and analyzed with Fiji (ImageJ-based software, National Institutes of Health, USA). For each sample, five sections were analyzed, with five regions of interest per section. The region of interest was defined as a full-field image acquired at 40× magnification. To avoid overlap, every fifth section was analyzed, ensuring a minimum spacing of 50 μm.
Statistical analysis
Obtained data was assessed for normality with Shapiro-Wilk test and unpaired Student’s t-tests were used for statistical analyses. Both statistics and graphs of the obtained results were performed using GraphPad Prism 10, version 10.5.0 for Windows, Academic License (GraphPad Software, USA).
RESULTS
Immunohistochemical staining showed distinct distribution patterns of the analyzed markers in control and diabetic epidermis.
In control samples, RAGE immunoreactivity was observed mainly in the granular layer, and in diabetic samples it was present in both the granular and basal layers. CML staining was detected across the epidermis in both groups, although in diabetic samples it appeared more intense and was more prominent in the basal layer.
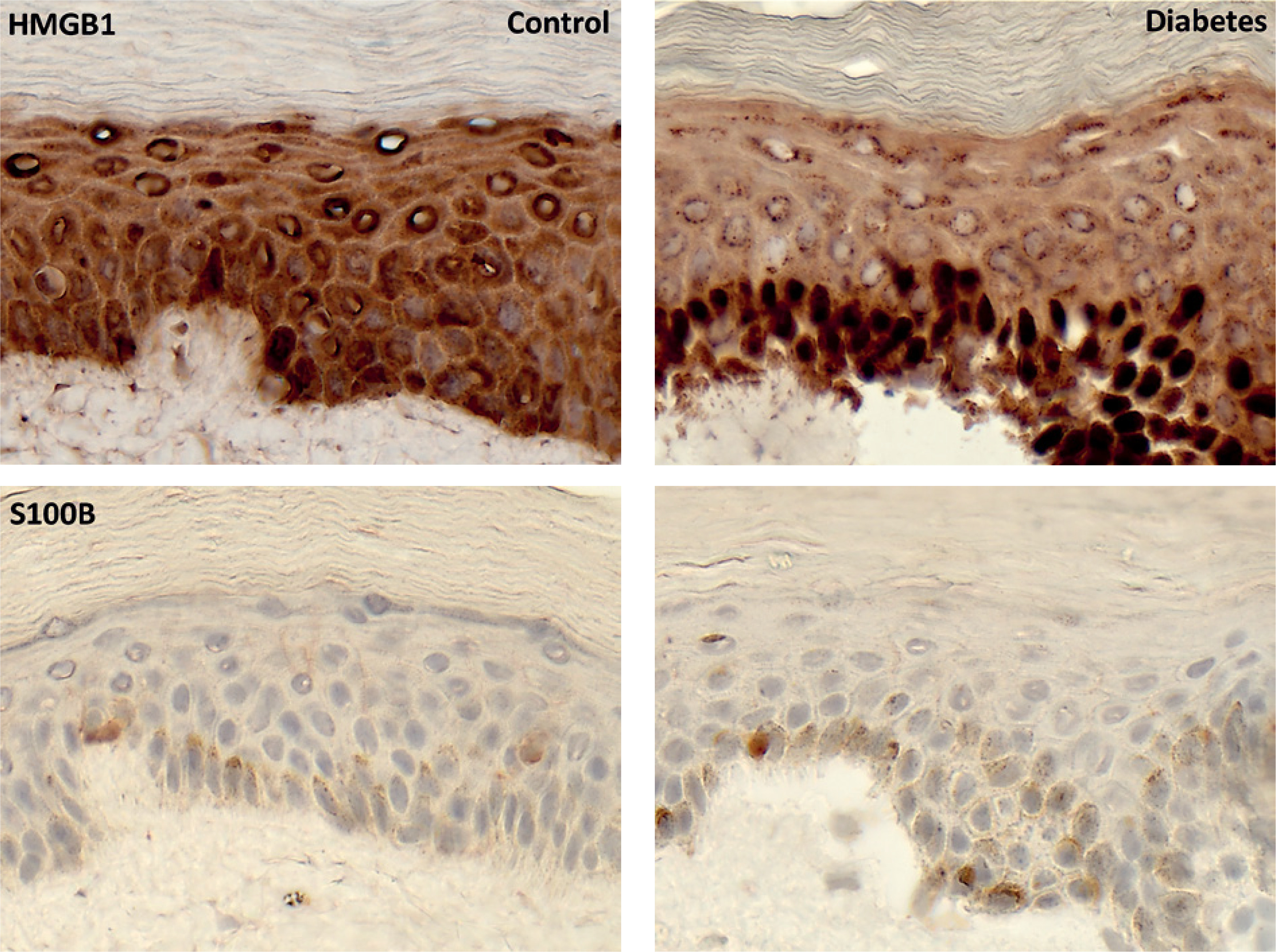
HMGB1 immunoreactivity was observed predominantly in the basal and lower spinous layers, with stronger staining in diabetic samples. S100B-positive staining was limited to individual epidermal cells in both groups, with a greater number of positive cells observed in diabetic samples (figs. 1, 2).
Figure 1
Immunohistochemical diaminobenzidine (DAB) staining of the receptor for advanced glycation end products and carboxymethyllysine. Brown deposits indicate receptor- or ligand-positive intra- and extracellular material surrounding epidermal cells. Sections were counterstained with hematoxylin. Differences in staining intensity and distribution are observed between control and diabetic epidermis. For epidermal layer identification, see figure 3. Magnification, ×400
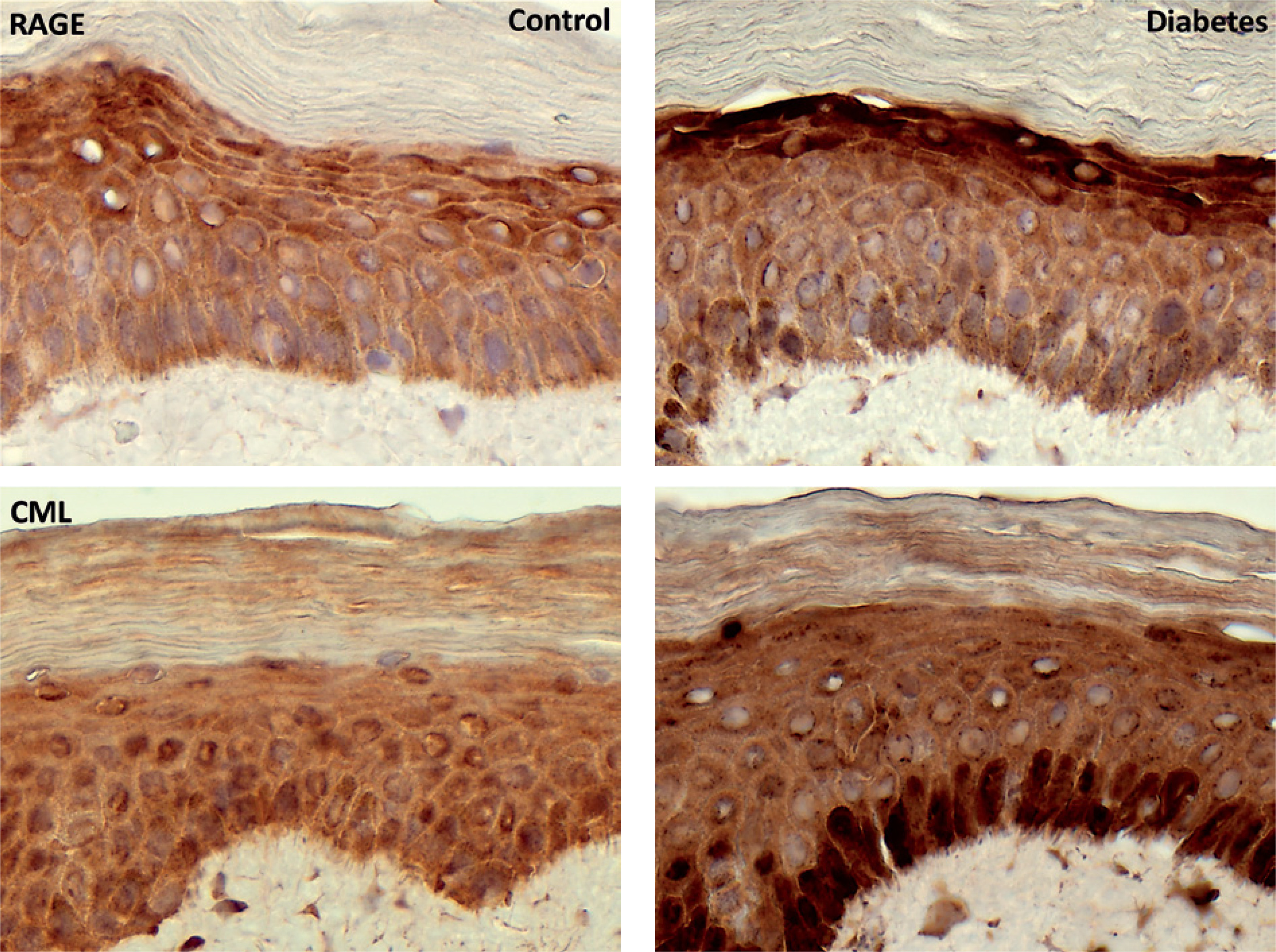
Figure 2
Immunohistochemical diaminobenzidine (DAB) staining of high-mobility group box 1 and S100 calcium-binding protein B. Brown deposits indicate positive intra- and extracellular material surrounding epidermal cells. Sections were counterstained with hematoxylin. Increased staining intensity for high-mobility group box 1 and a higher number of S100 calcium-binding protein B–positive cells are observed in diabetic epidermis. For epidermal layer identification, see figure 3. Magnification, ×400
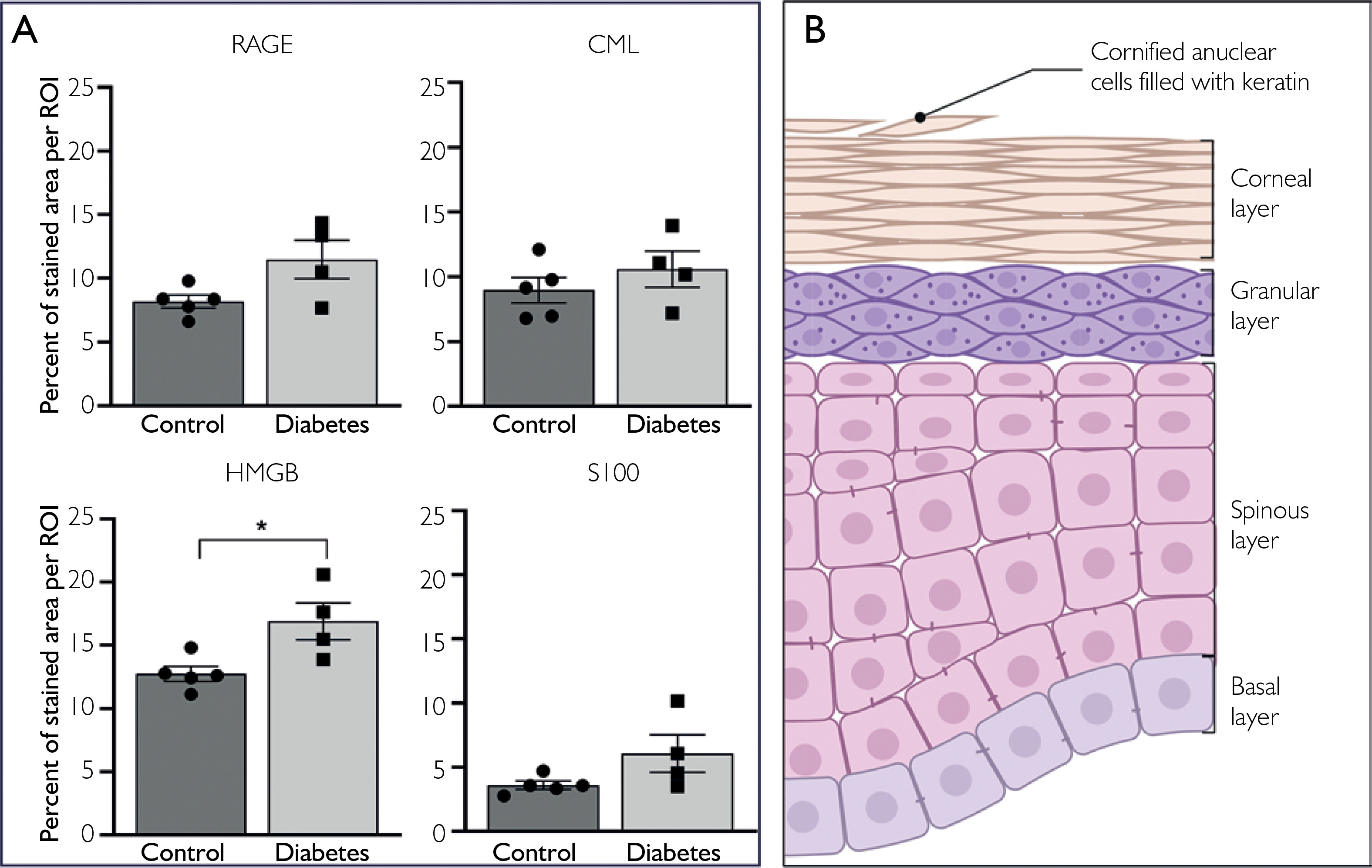
Semiquantitative analysis of stained area fraction showed numerically higher values in diabetic samples for all analyzed markers. For RAGE, the stained area fraction was 8.18 ±0.51 SEM in controls and 11.48 ±1.51 SEM in diabetic samples. For CML, the corresponding values were 9.96 ±0.98 SEM and 10.60 ±1.39 SEM. HMGB1 showed higher values than RAGE and CML in both groups and was the only marker for which the between-group difference reached statistical significance, increasing from 12.76 ±0.59 SEM in controls to 16.91 ±1.45 SEM in diabetic samples, p ≤ 0.05. S100B showed the lowest stained area fraction among the analyzed markers in both groups, with values of 3.60 ±0.31 SEM in controls and 6.07 ±1.45 in diabetic samples (fig. 3 A).
Figure 3
Statistical analysis and schematic representation of epidermal layers. A – Graph columns represent the percentage of stained area per region of interest (ROI), defined as a full-field image acquired at ×40 magnification. Data points represent individual samples. Higher values were observed for all analyzed markers in diabetic samples; however, statistical significance was reached only for high-mobility group box 1 (p ≤ 0.05; control, n = 5; diabetes, n = 4). B – Schematic representation of epidermal layers for reference. Created with BioRender
DISCUSSION
Our studies have shown that RAGE and its proinflammatory ligands are present in susceptible tissues in diabetes and its complications [12, 15]. Increased expression of RAGE and its ligands has been reported in diabetic kidney disease, atherosclerosis, diabetic peripheral neuropathy, and diabetic retinopathy [21–23]; however, their role in diabetic skin pathology remains insufficiently explored. Our current findings are consistent with our previous investigations, suggesting altered staining patterns of RAGE and its proinflammatory ligands in the epidermis of patients with diabetes, further supporting the involvement of RAGE in the pathogenesis of diabetic complications.
Evidence from human and animal studies associates RAGE with several pathological skin conditions, pointing to possible participation of RAGE in inflammatory responses relevant to tissue injury in diabetes [24]. A study by Park et al. showed that RAGE is present in endothelium of dermal and subcutaneous vessels in patients with severe diabetic neuropathy [25]. In addition, early studies demonstrated that RAGE-encoding gene (AGER) G82S polymorphism is associated with diabetic skin disease in patients with type 2 diabetes [26]. Furthermore, studies show that CML, a classical representative of AGEs and one of the primary ligands of RAGE, accumulates physiologically in aging skin, and this process is accelerated in diabetes [27]. A recently published study demonstrated that skin CML levels correlate with increased skin rigidity in the reticular dermis in diabetic mice [28]. Elevated levels of AGEs, including CML, have been also observed in skin of patients with type 1 diabetes and correlate with disease duration [29]. Skin AGE assessment, measured as skin autofluorescence, has been also investigated as a prognostic tool for the development and progression of diabetic peripheral neuropathy [30].
S100B, which is physiologically present in selected skin cell populations, has also been reported to show altered expression in several inflammatory and neoplastic skin disorders [31]. Similarly, another member of the S100 family, S100A7, shows increased expression not only in hyperproliferative skin diseases such as psoriasis and atopic dermatitis, but also in skin cancers, including melanoma and squamous cell carcinoma [24, 32]. In addition, studies have demonstrated elevated expression of S100A7, S100A8, and S100B9 mRNA in venous leg ulcers in otherwise healthy individuals [33]. Likewise, a study on diabetic foot ulcers showed that S100A9 expression was significantly increased in both blood and chronic wound samples from diabetic patients compared to non-diabetic controls [34].
Finally, elevated levels of HMGB1 have been reported in severe acne, lichen planus, and systemic lupus erythematosus [35]. Increased HMGB1 expression often coincides with elevated RAGE levels in various skin pathologies, indicating their potential roles as biomarkers and therapeutic targets. Recent translational studies may reflect the utility of HMBG1 as a biomarker in systemic lupus, while RAGE and AGEs have been proposed as potential biomarkers in psoriasis and skin aging [36]. A recent evidence indicates that increased HMBG1 expression in diabetic skin wounds impairs fibroblast function and prolongs wound healing under hyperglycemic conditions [37].
Despite extensive evidence supporting the role of RAGE and its ligands in various skin diseases, their role as biomarkers in diabetes-associated skin pathologies remains insufficiently explored. Moreover, although targeting the RAGE-ligand axis has been investigated in conditions such as lichen planus, fibrosis, and systemic lupus, relatively little is known about its role in the pathogenesis of diabetic dermopathy [24]. Recent studies have explored small-molecule inhibitors targeting RAGE-ligand interactions in inflammatory disorders. Although these data are of interest, the present study was not designed to assess therapeutic implications, and any translational relevance to diabetic skin pathology remains speculative [38].
Our study provides preliminary evidence that components of the RAGE-ligand signaling axis are detectable in diabetic epidermis and may show altered expression patterns relative to controls. This study has several important limitations. First, the sample size was very small, which substantially limits statistical power and increases the risk of both false-negative and unstable estimates. Therefore, the present findings should be interpreted as exploratory and hypothesis-generating rather than confirmatory. Second, the restricted size and composition of the study groups limit the generalizability of the results. Third, the analysis was based on immunohistochemical assessment of a limited number of skin samples and regions of interest, which, although standardized, provides primarily descriptive and semiquantitative information. Finally, the study was not designed to establish mechanistic relationships between marker expression and the development of diabetic skin pathology.
Accordingly, larger studies are needed to confirm these observations and define their biological and clinical relevance.










