

Introduction
ORAI1 and STIM1 genes are essential for the store-operated Ca2+ entry (SOCE) mechanism used by immune cells to increase intracellular Ca2+ concentrations [1]. Expression of these genes in blood of patients with tuberculosis (TB) became the focus of our exploration. We expected some impairment in this specific regulatory mechanism of Ca2+ entry in patients with TB.
Tuberculosis is an infection characterized by the persistence of mycobacteria in macrophages and dendritic cells due to imperfect phagocytosis. The local secretion of interferon γ (IFN-γ), produced by type 1 T helper cells, and tumor necrosis factor α (TNF-α), primarily produced by macrophages, are the main factors enhancing the maturation of phagolysosomes, production of reactive oxygen (ROS), nitrogen species, and antimicrobial peptides in macrophages [2]. Activation of immune cells, production of cytokines and inflammation-restricting proteins (Fas-ligand, Foxp3), and production of ROS in phagosomes of phagocytes occur after an increase of intracellular Ca2+ concentration [3, 4]. Many studies [5-9] indicate that mycobacteria use different strategies to manipulate Ca2+ signaling in macrophages and dendritic cells to increase their lifetime and escape from the immune response.
There are two main channels providing Ca2+ influx into the immune cells: Ca2+ release-activated Ca2+ (CRAC) channels, which are activated by decreasing intracellular Ca2+ concentration, and voltage-gated Ca2+ channels (VGCC), which are opened by depolarization of the cytoplasmic membrane [5, 10]. The immune cells predominantly use CRAC channels of SOCE [1]. The prototypical CRAC channel is formed by ORAI1 protein (encoded by the ORAI1 gene) located on the cytoplasmic membrane. Its activation is provided by ligation of stromal interaction molecule 1 (STIM1, encoded by the STIM1 gene) located on the membrane of the endoplasmic reticulum [10].
The importance of STIM1 and ORAI1 genes in TB was demonstrated in an experimental model in TB infected mice [3]. A defect of the STIM1 gene led to the death of mice due to pulmonary hyper-inflammation and loss of respiratory function. It suggests that a dysfunction of SOCE may contribute to imperfect mycobacterial phagocytosis and provide a basis for mycobacterial persistence. Simultaneously, there are no data about the expression of ORAI1 and STIM1 genes in humans with TB. Detection of the expression level of ORAI1 and STIM1 genes in blood despite their non-specificity for TB can provide additional information for a TB pathogenesis, prognosis, and monitoring of TB treatment.
This study aimed to detect the level of expression of ORAI1 and STIM1 genes in blood of TB patients at the beginning of anti-TB treatment in association with immunological status, assessed by levels of IFN-γ and interleukin 18 (IL-18) in serum, and to compare the expression of ORAI1 and STIM1 with a healthy control group. IL-18 was selected for the assessment of the immunological status as a pro-inflammatory cytokine, produced predominantly by activated monocytes/macrophages and having broad immunological effects [11], including an increase of production of IFN-γ by T cells and enhancing the protective Th1 immune response against mycobacteria. Simultaneously, increasing activity of IL-18 induces activation of nuclear factor-kB and expression of Fas ligands, which can lead to tissue destruction [11, 12]. An increasing level of IL-18 in serum also has a negative predictive value for HIV-associated TB [13].
Material and methods
The permission of the Ethics Committee of Riga East University Hospital (No. 9-A/17) was obtained for the study. The study included 45 patients, aged 20 to 86 years (mean age = 49.7 years, SD = 16.0, 33% females), with confirmed TB. The patients underwent observation and treatment from October 2017 to December 2019 in the Lung Disease and Tuberculosis Ward of Daugavpils Regional Hospital (Latvia). Inclusion criteria were: age of 18 and older, bacteriologically confirmed pulmonary TB, and drug-sensitive TB. Exclusion criteria were: age < 18, exclusively extrapulmonary TB, pulmonary TB with concurrent extrapulmonary TB involvement, pregnancy, imprisonment, mental disorders, and HIV-positive status. All patients had bilateral drug-sensitive tuberculous pneumonia and used first-line anti-tuberculous drugs according to the standard regimen.
The control group included 35 volunteers, aged from 21 to 73 years (mean age = 36.8 years, SD = 16.0, 30% females), without active TB and human immunodeficiency virus (HIV) infection. For the control group, the inclusion criteria were age of 18 and older, self-reported physical health, and no ongoing infection.
For analysis of ORAI1 and STIM1 expression, 1 ml of peripheral blood with EDTA was collected and stored at –20oC before detection. RNA was extracted from stored blood samples using the innuPREP Blood RNA Life Science Kits & Assays (Analytik Jena Company, Germany) according to the manufacturer’s procedures. The quality and quantity of extracted RNA were detected by spectrophotometry using the Nanofotometr NF80 (Implen GmbH, München, Germany). The concentration of RNA was 40 µg/ml per reaction. For reverse transcription-PCR (RT-qPCR), the Revert First-Strand cDNA synthesis system with an oligo dT primer (QuantiTect Reverse Transcription, an oligo dT primer Invitrogen, Germany) was used following the manufacturer’s instructions. The first-strand cDNA was diluted 1 : 20 with distilled water with the following used as a template (K1+) in RT-qPCR analysis. Specific primers for human STIM1 (Hs STIM1 FAM_1, QF00208159) and ORAI1 (Hs ORAI 1 FAM_1, QF00163611) were detected by one-step qRT-PCR using sequence-specific probes for gene expression analysis (QuantiFast Probe Assay, Invitrogen, Germany). RT-qPCR was performed in a DTlite Cycler (DNA-Technology, Russia) by the fast real-time PCR System using the following amplification conditions: 5 min of initial denaturation at 95°C, then 45 cycles of 95°C for 30 s, 60°C for 30 s. The specificity of RT-qPCR products was confirmed by the analysis of a melting curve. Absolute quantification of STIM1 and ORAI1 gene expression was detected relative to a standard curve, automatically created by serial dilution of the reference gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH). For amplification, the following primers (Invitrogen by Thermo Fischer Scientific) and reference gene (Bioneer Corporation, Republic of Korea) were used: ORAI1, forward: 5'-CGTATCTAGAATGCATCCGGAGCC-3' reverse: 5'-CAGCCACTATGCCTAGGTCGACTAGC-3'; STIM1, forward: 5'-CCTCGGTACCATCCATGTTGTAGCA-3', reverse: 5'-GCGAAAGCTTACGCTAAAATGGTGTCT-3'; GAPDH, forward: 5'-CCACTCCTCCACCTTTGAC-3', reverse: 5'-ACCCTGTTGCTGTAGCCA-3'; Probe, 5'-TTGCCCTCAACGACCACTTTGTC-3'.
Detection of IFN-γ and IL-18 in serum was performed by the sandwich immunoassay method using the commercial kits IFA-BEST INF-γ (detection range: 0-1000 pg/ml, sensitivity: 2.0 pg/ml, normal values: < 15 pg/ml) and IFA-BEST IL-18 (detection range: 0-1000 pg/ml, sensitivity: 2.0 pg/ml, normal values: 90-260 pg/ml) according to the methodology of the manufacturer (Vector-Best, Russia).
Results and discussion
The Shapiro-Wilk test revealed significant deviance from the normal distribution in both groups of participants. Test values varied from 0.19 to 0.91 (p < 0.01 to p < 0.001). Observed non-normality of distributions led to the use of nonparametric statistics for a comparison of the control and TB groups. The results of the Mann-Whitney U-test (Table 1) indicated a significantly higher level of mRNA ORAI1 in the control group.
Table 1
Comparison of markers in tuberculosis (TB) and control groups
Simultaneously, there were no significant differences between groups in the level of mRNA STIM1. The level of IFN-γ and IL-18 were higher in the group of TB patients. Figure 1 presents the comparison between groups.
Fig. 1
Differences between groups in mRNA ORAI1, mRNA STIM1, IFN-γ and IL-18 (medians and their 95% CI are presented on a decimal logarithmic scale)
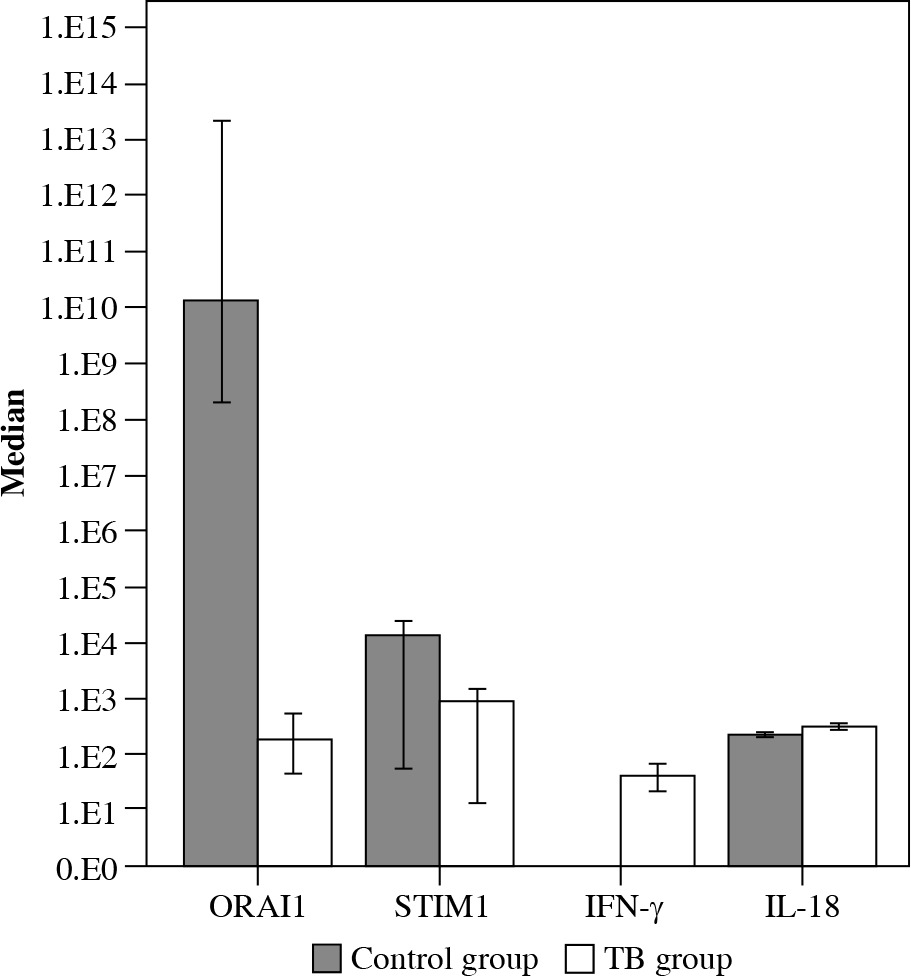
Expression levels of ORAI1 and STIM1 genes in blood dominantly reflected their expression in peripheral blood leukocytes, being immune cells. A more expressed ORAI1 gene in TB patients indicated an impaired SOCE mechanism in these cells.
Taking into account that ORAI1 protein forms a pore in the plasmatic membrane of immune cells for releasing Ca2+ ions into cytoplasm, a lower expression level of ORAI1 in blood can be associated with a lower influx of Ca2+ into cells, leading to lower cytokine production. In contrast, our findings showed that the production of IFN-γ and IL-18 is not disrupted in patients with TB and is at a higher level than in the control group. As described in a model of TB infected mice [3], impaired SOCE is associated with the reduced expression of inflammation-restricting proteins, leading to hyper-inflammation.
The Spearman correlation coefficients indicated no significant relationships among the markers in both the control and TB groups (Table 2). It indicates the relative independence of the level of expression of genes and the level of inflammation.
Table 2
Spearman correlations among markers in control and tuberculosis (TB) group
| Markers | Control group | TB group | ||||
|---|---|---|---|---|---|---|
| ORAI1 | STIM1 | IFN-γ | ORAI1 | STIM1 | IFN-γ | |
| STIM1 | 0.07a | – | 0.12c | – | ||
| IFN-γ | –0.18b | –0.38b | – | –0.10d | 0.16d | – |
| IL-18 | –0.05b | 0.06b | 0.13b | 0.15e | –0.03e | –0.06d |
Simultaneously, non-pronounced differences in STIM1 gene expression indicate a low association of its expression with TB. Ca2+ influx into immune cells is also provided by other Ca2+ channels [14], for example, VGCC [6]. In this case, Ca2+ influx through VGCC leads to suppression of the protective immune response [6, 7].
At a more generalized level, observed differences in expression of the ORAI1 gene have at least two explanations. One the one hand, low expression of the ORAI1 gene in patients with TB can be a result of mycobacterial infection. Previous studies [15-18] show that mycobacteria can change Ca2+ signaling and protect survival in macrophages by regulation of expression of microRNAs (endogenous regulators of gene expression) in TB patients. It is possible that microRNAs block ORAI1 expression. The relationship between ORAI1 expression and microRNA specific for TB can be the further direction of the investigation. On the other hand, we have not assessed the dynamics of ORAI1 expression. A low level of expression can be observed before the infection and affect the development of TB. A prospective study can present the dynamics of ORAI1 after the treatment. In addition, low expression of ORAI1 can be a result of genetic polymorphisms of the ORAI1 gene [19, 20].
Focusing on ORAI1 and STIM1 expression constitutes a limitation of the study because regulators of other Ca2+ channels and the intracellular level of Ca2+ were not included in the investigation. We assessed the expression of ORAI1 and STIM1 genes but did not directly address the level of expression of ORAI1 and STIM1 proteins. Moreover, assessing the expression of ORAI1 and STIM1 genes and proteins in blood should be performed in a larger group of patients with different severity of TB and should include monitoring of TB dynamics. Further studies should also include a broader range of Ca2+ channels and their regulators to investigate their complex functioning in TB patients.
Conclusions
It can be concluded that patients with TB have a lower level of ORAI1 expression in blood than individuals without TB. It indicates some impairment in the SOCE mechanism of immune cells, which is associated with TB. Despite the lack of significant correlations among ORAI1 and STIM1 gene expression levels and levels of two inflammatory cytokines, we have demonstrated that a low level of the ORAI1 gene and high levels of markers of inflammation are presented simultaneously in blood of TB patients.