Current issue
Archive
Manuscripts accepted
About the Journal
Editorial office
Editorial board
Abstracting and indexing
Subscription
Contact
Ethical standards and procedures
Most read articles
Instructions for authors
Article Processing Charge (APC)
Regulations of paying article processing charge (APC)
NEUROLOGY / CLINICAL RESEARCH
Klotho and vitamin D in multiple sclerosis: an Italian study
Marcello Ciaccio
1,3
1
Department of Biomedicine, Neuroscience, and Advanced Diagnostics, Institute of Clinical Biochemistry, Clinical Molecular Medicine, and Laboratory Medicine, University of Palermo, Palermo, Italy
2
Department of Experimental Biomedicine and Neuroscience, University of Palermo, Palermo, Italy
3
Department of Laboratory Medicine, University-Hospital, Palermo, Italy
Submission date: 2019-03-21
Final revision date: 2019-04-30
Acceptance date: 2019-05-01
Online publication date: 2019-08-02
Publication date: 2020-05-26
Arch Med Sci 2020;16(4):842-847
KEYWORDS
TOPICS
ABSTRACT
Introduction:
Low vitamin D levels have been recognised as an important risk factor for autoimmune diseases, including multiple sclerosis (MS). MS is a multifactorial disease, the pathogenesis of which contributes both to genetic and environmental factors. Polymorphisms in genes codifying molecules involved in vitamin D homeostasis have been associated with hypovitaminosis D. However, the influence of polymorphisms of Klotho, which codify a protein with a pivotal role in vitamin D metabolism, have never been investigated. The aim of this study was to evaluate the association among genetic variants of Klotho, namely rs1207568 and rs9536314, serum 25(OH)D3 levels, and multiple sclerosis (both risk and disease progression).
Material and methods:
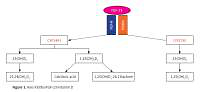
107 patients with MS and 133 healthy controls were enrolled in this study. Serum 25(OH)D3 levels and genotyping of Klotho SNPs were evaluated in all participants by high-performance liquid chromatography and real-time polymerase chain reaction, respectively.
Results:
Allelic and genotypic frequencies did not differ between patients and controls. Concerning rs1207568, we found a trend toward lower serum 25(OH)D3 levels in MS patients with A allele (mutant), both in heterozygosis (AG) and in homozygosis (AA), in comparison to MS patients with G allele in homozygosis (GG) (AG + AA 20.5 ±6.3 µg/l; GG 22.5 ±7.5 µg/l, p = 0.07).
Conclusions:
Our findings did not identify a role of Klotho in the genetic susceptibility to MS.
Low vitamin D levels have been recognised as an important risk factor for autoimmune diseases, including multiple sclerosis (MS). MS is a multifactorial disease, the pathogenesis of which contributes both to genetic and environmental factors. Polymorphisms in genes codifying molecules involved in vitamin D homeostasis have been associated with hypovitaminosis D. However, the influence of polymorphisms of Klotho, which codify a protein with a pivotal role in vitamin D metabolism, have never been investigated. The aim of this study was to evaluate the association among genetic variants of Klotho, namely rs1207568 and rs9536314, serum 25(OH)D3 levels, and multiple sclerosis (both risk and disease progression).
Material and methods:
107 patients with MS and 133 healthy controls were enrolled in this study. Serum 25(OH)D3 levels and genotyping of Klotho SNPs were evaluated in all participants by high-performance liquid chromatography and real-time polymerase chain reaction, respectively.
Results:
Allelic and genotypic frequencies did not differ between patients and controls. Concerning rs1207568, we found a trend toward lower serum 25(OH)D3 levels in MS patients with A allele (mutant), both in heterozygosis (AG) and in homozygosis (AA), in comparison to MS patients with G allele in homozygosis (GG) (AG + AA 20.5 ±6.3 µg/l; GG 22.5 ±7.5 µg/l, p = 0.07).
Conclusions:
Our findings did not identify a role of Klotho in the genetic susceptibility to MS.
Share
RELATED ARTICLE
| eISSN: | 1896-9151 |
| ISSN: | 1734-1922 |
We process personal data collected when visiting the website. The function of obtaining information about users and their behavior is carried out by voluntarily entered information in forms and saving cookies in end devices. Data, including cookies, are used to provide services, improve the user experience and to analyze the traffic in accordance with the Privacy policy. Data are also collected and processed by Google Analytics tool (more).
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.
You can change cookies settings in your browser. Restricted use of cookies in the browser configuration may affect some functionalities of the website.