
Introduction
The reported prevalence of atopic dermatitis (AD) in children and adults varies worldwide. For instance, the prevalence of AD is the highest in Swedish children (34%) and the lowest in Tunisian children (0.65%). In the adult population, available data from 2021 suggest that AD occurs in 2–5% of this group [1].
Currently, the diagnosis of AD is mainly based on the clinical symptoms and criteria of Hanifin and Rajka [2]. Although they have been well known and practiced for many years, they are still insufficient in many cases. Therefore, biomarkers of disease have been searched for many years to help in predicting the risk of the disease, facilitating diagnostic processes, and increasing the efficacy of treatment. The clinical symptoms of AD may be simple, such as localized eczema, dry and altered skin, or pruritus. They are localized mainly in the flexures and can embrace the entire body. In infants, it is typically located on the extensor side. Moreover, AD is often associated with more complicated aspects, a such as psychiatric and mental health disturbances, sleep disorders, and depression. Complex aetiology and pathophysiology consist of different elements, both epigenetic and genetic, immunological factors, skin barrier defects, microbiome dysbiosis and environmental interactions [3]. The above-mentioned aspects will be developed in more detail in the following sections.
Aetiology
AD aetiology is complex, multifactorial, and still not fully understood. The course of AD is highly variable and unpredictable. It involves both immunological disorders and skin barrier dysfunction. The concept of immunological aetiology is still evolving. Recent studies support the activation of multiple Th-lymphocyte subsets leading to a full-blown disease. It is also connected with other inflammatory diseases sharing some similar mechanisms, such as atopic disorders: allergic rhinitis, asthma, food allergy and non-atopic comorbidities like cardiovascular diseases, neurological or psychiatric disorders connected with neuroinflammation, as well as autoimmune disorders [4]. Development of AD also depends on environmental and personal features and consists of skin barrier defects like dysregulation of cornified envelope proteins, anti-microbial peptides, lipids and tight junctions resulting in inappropriate pH, chronic inflammation within skin layers and increased transepidermal water loss and dry, itchy skin. Therefore, the microbiota is another example that gets disturbed following poor physiological microflora diversity and susceptibility to colonization by more virulent strains like Staphylococcus aureus, triggering an inflammation process [5]. Yet another factor causing AD is an immunologic disorder involving Th2, Th22, Th17 and Th1 pathways. It is widely developed, but still unknown to the end. This is well illustrated in the example of the treatment with modern drugs such as biologics and JAK inhibitors. They present different approaches to therapeutic strategies treating AD as not only a local skin disease [6].
Extracellular matrix (ECM)
Skin plays an important role in maintaining overall human health. The ECM is now known to play a role in both intracellular interactions and the inflammatory process of affected tissues in response to pathological stimuli [7]. The outermost layer of the skin is composed of many layers of dead, exfoliated keratinocytes. Recent reports proved ECM can prevent or help with skin infections [8]. Based on the evidence collected in the study, despite the limited knowledge in this field so far, it was possible to establish that changes in the matrix may be helpful in neoplastic research and the prevention of metastasis or cancer treatment [9]. Inflammatory skin diseases and impaired healing can lead to ECM remodelling, excess collagen deposition, and skin fibrosis. Research on the potential link between ECM modifications and skin diseases such as psoriasis and AD may provide new treatment targets [10].
Epidermal messengers
Kallikreins (KLKs) are key proteins of the epidermal microenvironment, regulating its exfoliation and inflammation. Dysregulation of KLKs is implicated in the pathogenesis of skin inflammation. Besides, they can influence inflammation and constitute activators of the skin repair system. Degraded ECM homeodynamics may be responsible for ineffective tissue repair and fibrosis [11]. The analysis of diseases with delay in other organs shows that the epithelium plays a crucial role in the persistence of the disease, and recent reports also indicate the role of keratinocytes in the formation of scleroderma [12]. Considering the skin, KLKs can influence dermatitis and proteolytic activity on the ECM, which may cause pathological remodelling of the ECM [13].
Collagens
Until now, it has been shown that in elastic connective tissues such as the skin, lungs and vascular system, type I collagen (Col I) often co-occurs with type III collagen (Col III). Although the main types of collagen present in the extracellular matrix are collagen types I and III, collagen types IV, V, VI and VIII can also be found in the extracellular matrix [14]. Type I collagen creates a skeleton of thick fibres characterized by a low elasticity factor. The maturation process of type I collagen is closely dependent on type III collagen, which creates thin, less durable fibres with greater elasticity [15]. The expression of collagen-encoding genes is regulated by the promoters of the Col I and Col III genes, which contain regulatory elements related to transforming growth factor β (TGF-β). Since the Col I gene promoter also contains specificity protein 1 (SP-1), binding sites missing in the Col III gene promoter, differences in the expression levels of the Col I and Col III genes may be due to differences in their promoters [16]. Collagen IV is the collagen that is initially present in the extracellular matrix basement membranes (BM) that separate various epithelial and endothelial cells. It is an important component of the connection between the skin and the epidermis.
Type IV collagen molecules form complex protein networks that are a major component of the basement membrane, thin sheet-like structures that separate and support cells in many tissues [17].
Differences in the levels of Col I and Col III may result not only from differences in gene expression but also from those in the degradation process of these proteins. Two different matrix metalloproteinases (MMPs) involved in collagen degradation have been identified. MMP-1, produced by fibroblasts, has an equal tendency to degrade both Col I and Col III. However, MMP-8, produced by neutrophils, has a greater predisposition to the degradation of Col III. Therefore, differences in MMP activity or expression may also influence changes in Col I and Col III content, in addition to changes in gene expression [18].
The key role of collagens in humans has been demonstrated by the variety of illnesses caused by genes encoding them [19]. Some studies showed a connection between epidermic collagen levels and manifestations of allergic disease. In 2007, scientists proved a lack of COL29A1 gene expression is a feature of AD, showing polymorphisms in genes encoding collagens that may generate susceptibility to such illnesses as AD [20]. Another study looked at collagen tripeptide (CTP). Researchers showed that in human keratinocytes mRNA and protein chemokine levels were significantly inhibited by CTP treatment in AD-like inflammatory conditions. It has been observed that CTP may provide therapeutic benefits in AD by inhibiting allergic type 2 inflammation [21]. Another study investigated the COL6A5, COL8A1 and COL10A1 genes as potential AD susceptibility biomarkers. The transcript level of COL6A6 has been shown to be higher in people with AD than in the control group. In the tissues of AD patients, the transcript level of COL6A6 was initially lower in the epidermis compared to the control group, and the level of COL6A6 increased in the dermis. It was noticed that a decrease in the level of COL6A6 in the epidermis may indicate damage to the skin barrier and contribute to the early onset of atopic dermatitis [22, 23]. The results of one study showed that the COL8A1 gene may be a new susceptibility factor, increasing the incidence of atopic eczema (AE). However, the study was limited by its small sample size [24]. Despite the above-gathered data from published studies, little is known about the exact contribution of the different types of collagens present in the human skin to AD. However, the above-mentioned studies suggest an interesting perspective on the origins of AD.
Aim
The primary objective of our research has been to determine the mRNA levels of several collagens located in non-lesional and lesional parts of the skin in patients suffering from AD and their correlation to healthy subjects. In addition, we conducted a brief clinical evaluation of the study group of people with AD in terms of the severity of the disease evaluated with the SCORAD scale and the pruritus intensity measured using the VAS scale. Finally, we wanted to compare the obtained expression results for individual collagens with the clinical assessment of patients with AD.
Material and methods
Patients and sample
The study group consisted of 9 AD patients (men and women, aged 21–64, median age 40) who were patients of the Department of Dermatology, Venereology and Allergology of the Medical University of Gdansk/University Clinical Centre. Patients with no comorbidities nor topical treatment for at least 14 days except for emollient therapy were enrolled in the study. The control group included 9 healthy people (men and women, aged 21–64, median age: 53) without any inflammatory, malignant disease or allergies and not using topical or systemic glucocorticoids who were patients of the Plastic Surgery Clinic of the Medical University of Gdansk/University Clinical Centre. AD was diagnosed based on the criteria of Hanifin and Rajka [2]. Punch biopsies for analysis of collagen gene expression were obtained from 9 AD patients (9 from lesional sites and 9 from non-lesional sites) and 9 healthy controls (9 from non-lesional healthy skin). The collection sites were similar and concerned with the intragluteal fissure and the iliac crest. The biopsies were 4 mm in length and 0.7 mm in depth. 6 out of 9 samples from the AD group were good enough to be tested for collagen expression. The study was approved by the Ethics Committee of the Medical University of Gdansk (NKBB/71/2020) and we conducted it in accordance with the principles of the Declaration of Helsinki. All volunteers participating in the study signed written consent before being included in all the study procedures.
Isolation of total mRNA
Total RNA was isolated by the method of Chomczynski with our modifications [24]. The skin fragments were 4-mm punch biopsies homogenized in a sterile tube of 1 ml of TRI reagent (Sigma-Aldrich, Poznañ, Poland). Next, we added chloroform (250 μl), shook the samples, incubated them at 4°C for 15 min and centrifuged (10,000× g for 15 min at 4°C). The upper aqueous phase was removed into a new Eppendorf tube and an equal volume of isopropanol was added. RNA was precipitated by overnight incubation at -20°C followed by centrifugation (10,000× g for 15 min at 4°C). Subsequently, RNA pellets were washed with 96%, which was followed by 75% (v/v) ethanol, air-dried, resolved in diethyl-pyrocarbonate-treated thermo-sterilized water (15 μl) and stored at -20°C until further analysis.
mRNA level determination
The mRNA levels of COL1A1, COL2A1, COL3A1, COL4A1 genes were determined by real-time RT-PCR with appropriate TaqMan Universal Probe Library set (Roche Applied Science) using the Path-IDTM Multiplex One-Step RT-PCR kit (Path-ID Multiplex One-Step RT-PCR Kit, Applied Biosystems, Foster City, CA, USA). The levels of the tested transcript were normalized to the transcript level of the β-actin reference gene (ACTB). The reaction was performed with a final volume of 10 μl of the reaction mixture, containing: 1× Multiplex RT-PCR buffer, 1 μl of Multiplex enzyme mix, 0.5 μM of each PCR primer, 0.2 μM of each hybridization probe, and approximately 10 ng of total RNA isolated from the analysed tissue. The data analysis was performed using the LightCycler 480 II software (Roche Diagnostics International Ltd, Rotkreuz, Switzerland). The primer sequences, TaqMan probe and cycling conditions used are listed in Table 1 [25].
Table 1
The primer sequences, TaqMan probe and cycling conditions
Statistical analysis
The statistical analysis was conducted by using GraphPad Prism5. A comparison between lesional and non-lesional changed skin within the same subject was performed with the paired t-tests and the Wilcoxon signed ranked test. Afterwards, we compared the results from the study group of patients with AD to the control group of healthy subjects using the Mann-Whitney test. The graphic analysis was performed with the use of GraphPad Prism5.
Results
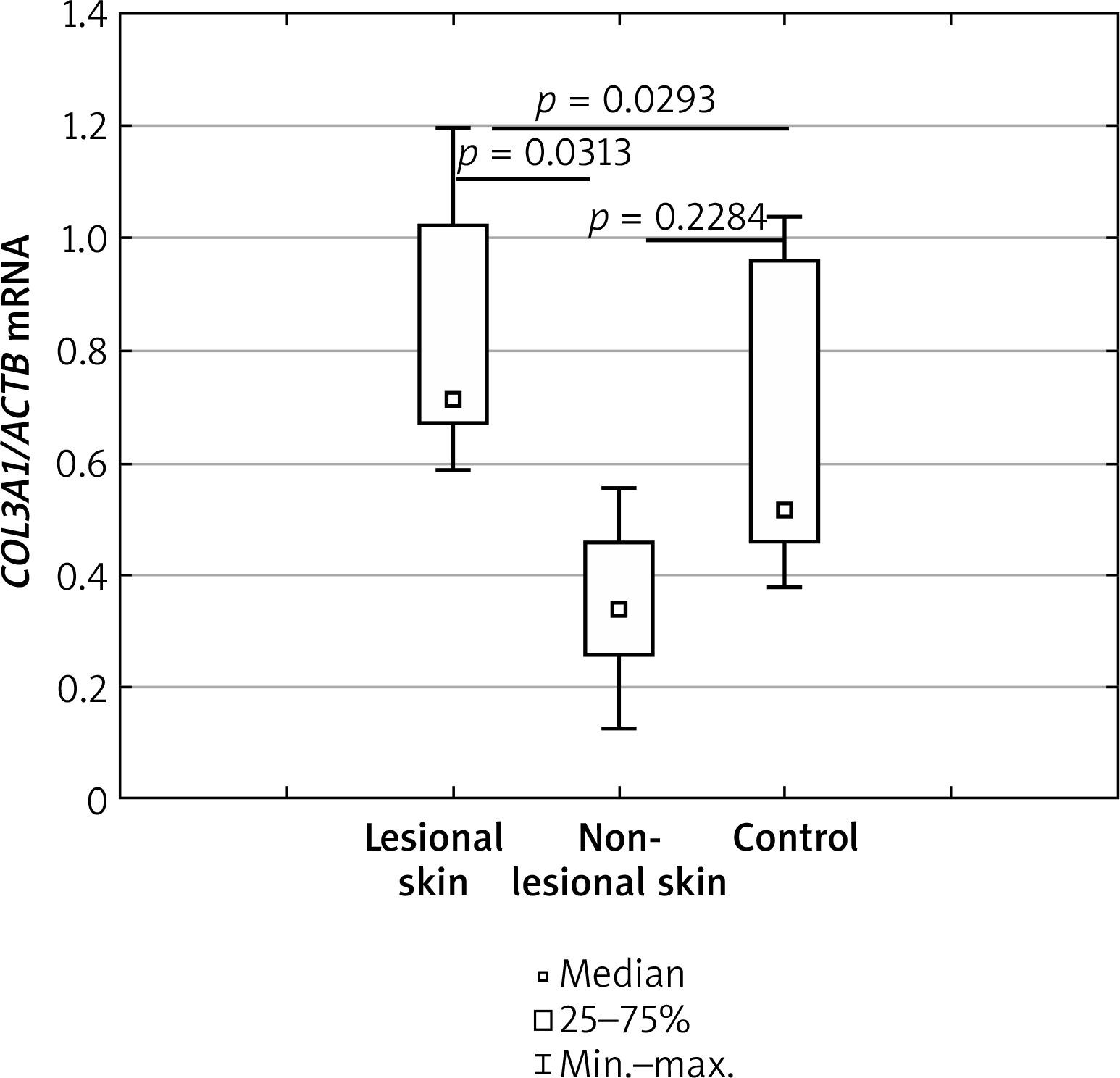
mRNA level of COL3A1 gene
The mRNA level of the COL3A1 gene was significantly higher in lesional skin than in non-lesional skin of AD patients in the study group (p = 0.0313). Compared to healthy controls, COL3A1 mRNA was significantly higher in the lesional skin of AD patients (p = 0.0293), however, there was no significant statistical difference between the non-lesional skin of AD patients and healthy controls (p = 0.2284). Details have been shown in Figure 1.
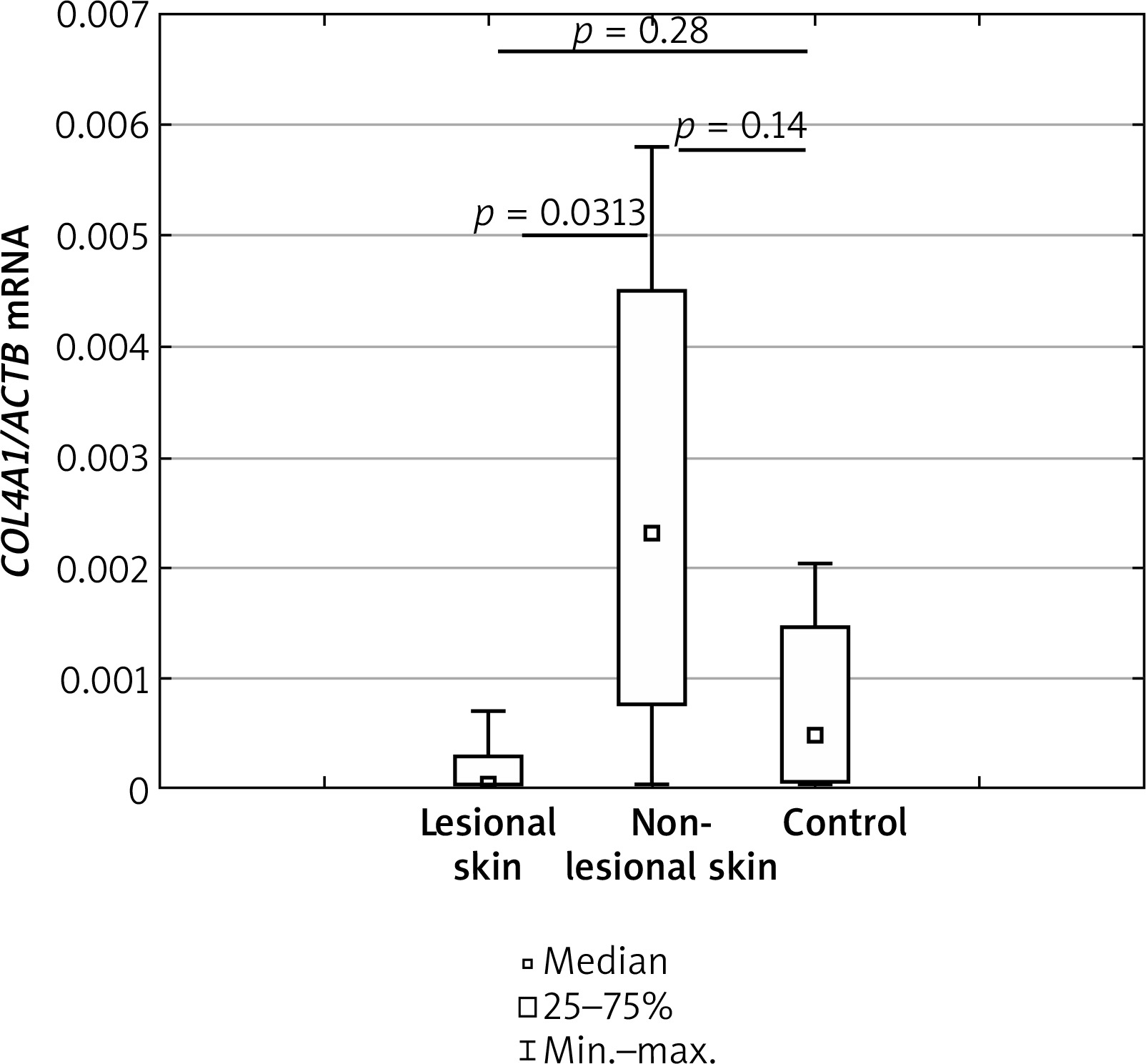
mRNA level of COL4A1 gene
The transcript level of COL4A1 was significantly higher in the AD skin from non-lesional sites than from the lesional sites (p = 0.0313). Details have been shown in Figure 2. The expression levels of mRNA COL4A1, when compared in lesional and non-lesional skin of AD with healthy controls, showed no statistically significant differences (p = 0.28 and p = 0.14, respectively).
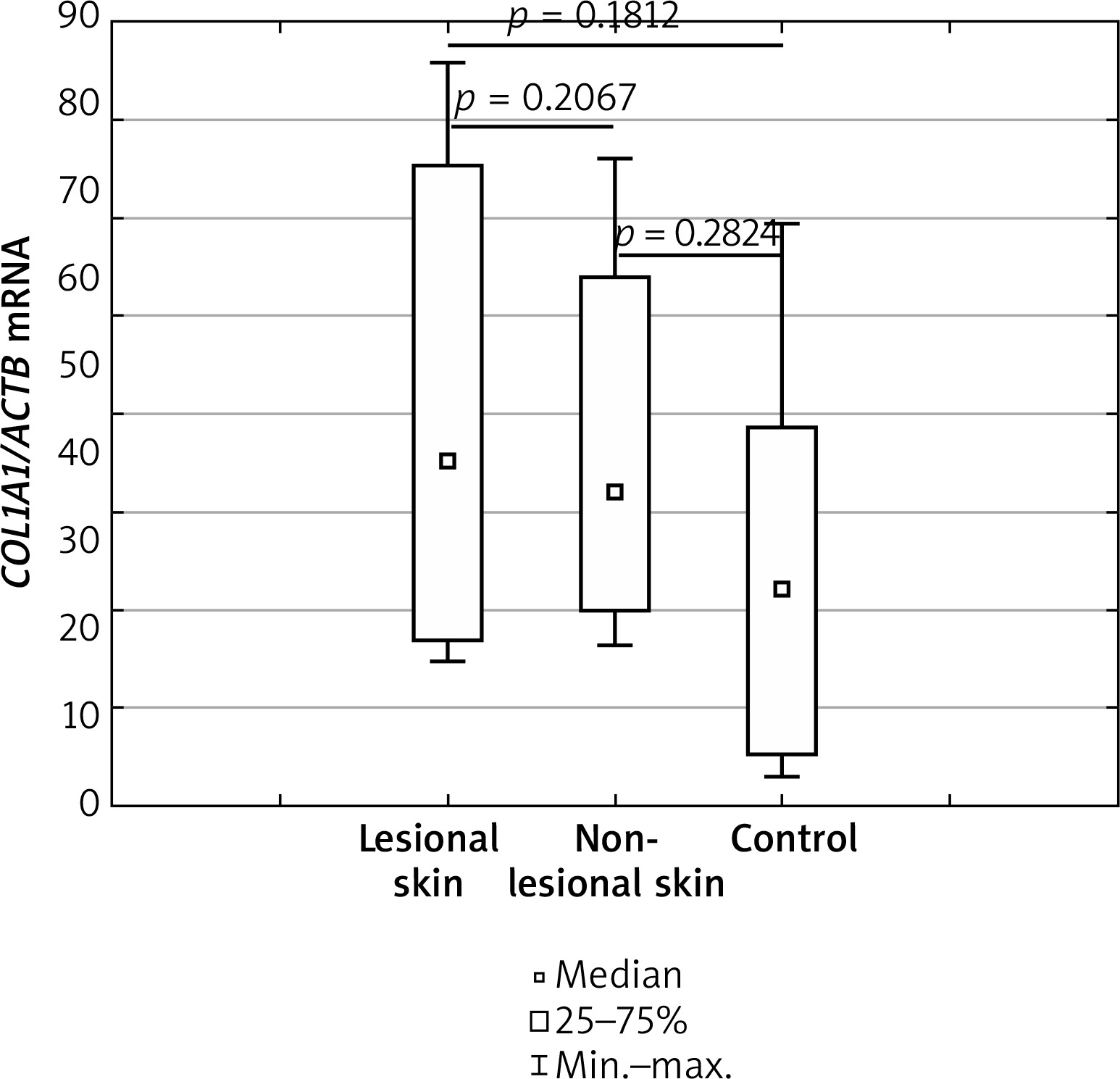
mRNA level of COL1A1 gene
Regarding the mRNA levels of COL1A1, no significant differences were observed between AD skin from non-lesional and lesional sites (p = 0.2067). In addition, no statistically significant differences were obtained after comparing expression levels of the COL1A1 gene in lesional and non-lesional skin of AD with healthy controls (p = 0.1812 and p = 0.2824, respectively). Details have been shown in Figure 3.
mRNA level of COL2A1 gene
No expression of the gene encoding type II collagen (COL2A1) was found in the skin.
Correlation between clinical symptoms, AD severity and the mRNA level of genes encoding type I, III and IV collagens
In the group of AD subjects, a higher level of COL3A1 mRNA expression correlated with a higher VAS score of pruritus. Moreover, lower levels of mRNA were associated with a decrease in pruritus severity using the VAS scale. It should be assumed that the intensity of itching is subjective and strongly limited by individual and external factors that may modulate its sensation. The above relationship has not been observed in the correlation between the level of COL4A1 mRNA expression and the intensity of the pruritus described in the VAS scale. Therefore, the results in question appeared to be coincidental. A similar relationship has been obtained to describe the intensity of the disease measured by the SCORAD scale and the transcript level of COL3A1. Higher levels of mRNA COL3A1 gene expression were noted for severe disease, which has not been observed for mRNA COL4A1 gene levels, thus displaying a random distribution of the examined feature. The study of the association of clinical symptoms with collagen mRNA expression levels was not statistically significant (p > 0.05).
Discussion
The collagen network constitutes a dynamic metabolic structure and collagen turnover, which most likely occurs between days 80 and 120, is regulated by the balance between collagen production and degradation [26–34]. The fibroblasts and myofibroblasts that are involved in this process respond to a variety of factors, such as mechanical stress, locally produced autocrine and paracrine substances (e.g. TGF-β and growth factors such as angiotensin II) as well as hormones supplied from the circulation, such as aldosterone. Additionally, various pro-inflammatory cytokines secreted by monocytes and macrophages influence the activity of fibroblasts and myofibroblasts [35]. The process of resolution of inflammation is dynamic and depends on the balance between pro-inflammatory and anti-inflammatory reactions. According to one study, the use of a stabilized collagen matrix induces an intense and short-lived inflammatory response that promotes rapid wound healing [36]. In addition, collagen improves the mechanical strength and elasticity of the tissue and serves as a natural substrate to support cell adhesion, proliferation and differentiation. MicroRNA-mediated regulation of MMP-2 at the wound site, as well as through interactions with the biofilm, may create an environment conducive to collagen degradation. This in turn leads to a reduction in the collagen I to collagen III ratio and a deterioration in the biomechanical properties of the repaired skin, potentially increasing the risk of wound recurrence [37].
Several studies have shown that major changes in collagen protein composition are typically characterised by a significant change in the ratio of type I to type III collagen [38]. From a pathophysiological point of view, mature type I collagen is primarily responsible for mechanical stability, whereas type III collagen, which forms thin bands, is often considered to be the juvenile collagen present in the early stages of wound healing [39].
Scientific reports indicate that collagens, the major component of the extracellular matrix (ECM), are essential for maintaining the integrity and proper function of the epidermis [5, 7, 8].
In turn, numerous reports highlight dysfunction of epidermal barrier integrity as a key factor in the development of AD [22–23]. It is also known that ECM collagens influence the migration of Th lymphocytes and epidermal antigen-presenting Langerhans cells and may therefore be involved in the regulation of the inflammatory response in the skin [23, 24]. In addition, collagens have been shown to bind to inhibitory immune receptors such as LAIR-1 on peripheral mononuclear cells, including NK, T and B cells. Down-regulation of ligands, which may be collagens, for these receptors can lead to over-activation of the immune system, resulting in chronic inflammation. This fact makes collagens particularly interesting targets in the context of AD disease [25, 26]. It has been reported that there is an association between some types of collagens and susceptibility to AD. The 2007 study identified genetic markers of subregions associated with AD. The region identified was COL29A1, the expression of which was absent in the outer epidermis of AD patients [5]. Several studies have suggested that risk factors for the development of AD lie in the genes that code for collagen [18]. The study showed a possible association between three polymorphisms located in the COL6A, COL8A1 and COL10A1 genes and potential susceptibility indicators for AD in a Mediterranean population.
In our study, we decided to investigate the expression levels of collagen type I, II, III and IV in the skin of AD patients compared to the normal skin of healthy individuals. Our results indicate a possible impact of the disease process of AD on the level of collagen type III and IV expression. In the samples tested from lesional sites, we observed an approximately twofold higher expression level of mRNA encoding collagen type III compared to its expression in non-lesional sites. In addition, the lesional skin of AD showed a significantly higher level of mRNA expression of this type of collagen compared to the skin of healthy individuals. Collagen III is known to be involved in wound healing. It is the first of all collagen types to be synthesised in the early stages of this process [27]. Thus, the observed overexpression of collagen III in the lesional skin of AD may be a response to skin injury caused by scratching. However, further research is needed to investigate the precise role of collagen type III in the pathophysiology of AD. Regarding the expression of collagen IV, we observed its lower expression in skin from lesional sites of AD compared to non-lesional sites. Perhaps the inflammatory process in active AD lesions is a cause of the lower expression of collagen type IV in these sites. IL-13, which contributes to the development of inflammation in AD, has also been shown to stimulate keratinocytes to produce matrix metalloproteinase-9, which can degrade type IV collagen [28]. Insufficient amounts of type IV collagen, which is the major collagen component of the basement membrane component – a structure important for the physical support of the epithelium – can compromise the structural integrity of the skin barrier [29]. This in turn can facilitate further penetration of harmful factors or allergens throughout the skin, leading to an inflammatory response and creating a vicious circle.
Overall, our findings of reduced collagen IV expression in lesional skin may indicate that the processes responsible for the development of AD lead to a reduction in collagen type IV expression. With regard to collagen III, its increased expression in lesional skin may also serve as a new biomarker for AD. In conclusion, our study provides data linking differences in collagen expression with the presence of AD. These results suggest that collagen III and IV are potentially interesting new biomarkers for AD. The obtained and observed differences in mRNA levels may be a result of deeper and more profound genetic changes in skin with developed AD. However, further studies with large groups of patients are needed to draw a more accurate and insightful conclusion. The more we know about skin barrier disorders, the better we can treat AD patients and prevent the development of atopic disease.
Conclusions
Our results indicate that there is a statistically significant difference between the expression level of genes encoding type III and IV in lesional and non-lesional parts of the skin and between the subject group and healthy individuals, especially for the mRNA of the COL3A1 gene. Further research is needed to evaluate the role of COL3A1 as a predictor of AD and its intensity or as a new indicator of AD severity. In addition, the expression levels for COL4A1 mRNA may be an indicator of the severity of epidermal barrier damage or an indicator of inflammation. In addition, it was confirmed that the expression of mRNA COL2A1 does not occur in the dorsal area around the iliac crest and the intragluteal fissure in both healthy volunteers and AD patients. A better understanding of its pathogenetic background, deep molecular mechanisms at the lowest level as DNA, could allow us to create new schemes for more reliable diagnosis and prediction of AD. The results of our study are particularly important in the light of personalized medicine. This approach follows the current trend of individualized treatment of patients. A better understanding of the underlying mechanisms of skin barrier dysfunction and immunological responses may allow better management of patients suffering from AD.







