







Introduction
The global demand for food is increasing in tandem with the growth of the human population. Agriculture is the primary source of our food supply, but its efficiency is closely linked to the climate. Climate change leads to long-term and seasonal variations in weather conditions, waterlogging, freshwater availability, increases in air temperature, and declines in relative humidity, as well as extreme rainfall events and seawater intrusion in arid lands (Miron et al., 2023). Agricultural lands located near coastal areas are highly diverse, fragile, and vulnerable. Furthermore, high rainfall and floods during the wet season, soil salinization during the dry season, acidity, high organic matter, and nutritional toxicities or deficiencies throughout the year contribute to low and unstable agricultural productivity. According to Mukhopadhyay et al. (2021), over 954 million hectares of land in 120 countries are affected by salt, with these salt-affected soils covering 20% of cultivated and 33% of irrigated lands, and contributing to an 8% loss of productivity. According to Mukhopadhyay et al. (2021), the rate of increase in salt-affected areas is expected to accelerate by 2050. Salinization would affect approximately 600 million people living in the coastal zone, who are primarily involved in agriculture and aquaculture activities (Ismail et al., 2022).
Changes in weather conditions and the emergence of new diseases can have adverse effects on agricultural growth and production (Corwin, 2020). Over the past two decades, newly developed rice varieties that are tolerant to salinity and/or floods have been deployed in affected areas, resulting in remarkable impacts (Bhowmick et al., 2020). These varieties have provided opportunities for designing better stress- and variety-specific management options and have given farmers more confidence to invest in input use and good crop husbandry (Imhoff-Kunsch et al., 2019; Bhowmick et al., 2020).
Enhancing the productivity and profitability of ricebased cropping systems in these coastal areas, with assured quality management services through proper harvest, postharvest processing, and value addition, will significantly improve smallholder farmers' livelihoods. Significant capital investment, large-scale adaptations to relevant technologies, and knowledge of know-how have demonstrated positive impacts in managing floods and salt intrusion, and in maximizing the agricultural lands in the potential coastal zone for agri-food production. Site-specific reclamation management strategies such as amendments, irrigation and drainage, and the use of environmental tolerance of planting materials and microbes are the components of climate-smart agriculture that can ensure food security (Jat et al., 2019; Bhowmick et al., 2020). Figure 1 illustrates the potential application of microbes in food production.
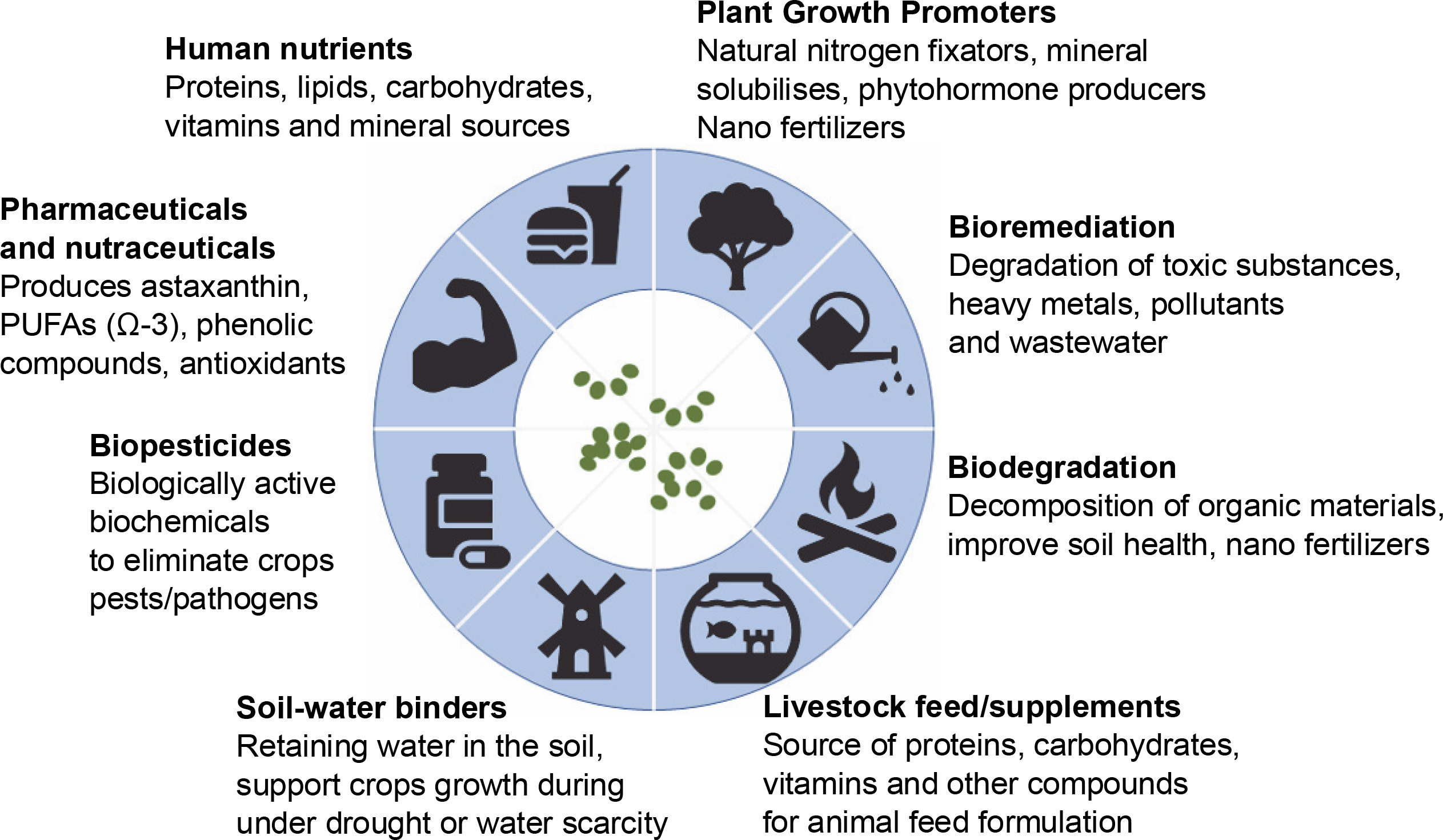
Foods and nutrient sources for humans and livestock
Microbes can be manipulated and used as food sources and nutrient providers for humans and livestock. Microbes can produce carbohydrates, lipids, proteins, minerals, and vitamins (Wang et al., 2021; Akkaya and Can-Guven, 2022; Kurniawan et al., 2022). Proteins are essential in human diets and animal feed formulations (Silva et al., 2022; Pyne et al., 2022). Aspergillus, Corynebacterium, and Brevibacterium are examples of amino acid sources used in dietary and feed supplements (Aizat et al., 2019; Karimi et al., 2021). Lysine, methionine, threonine, and tryptophan are commonly used as additives in animal feed formulations (Aizat et al., 2020). Moreover, mammals cannot synthesize vitamins and need to take dietary supplements to maintain balanced metabolism (Obeid et al., 2019). Cyanocobalamin (vitamin B12) can be produced through direct fermentation by Propionibacterium and Pseudomonas, whereas riboflavin (vitamin B2) can be produced from the fermentation of ascomycetes, Eremothecium, Bacillus, Corynebacterium, and Ashya (Obeid et al., 2019; Lordan, 2021). Additionally, organic acids such as lactic acid can be produced in the fermentation process performed by Aspergillus, Lactobacillus, Rhizopus, and Bacillus, and used in various foods and beverages (Tran and Zhao, 2021).
Microalgae, such as Chlorella, Spirulina, Aphanizomenon, Chlamydomonas, and Scenedesmus have become important microorganisms used as food and nutrient sources for humans and livestock worldwide (Barkia et al., 2019; Wang et al., 2021). Research has been conducted to increase the growth and production of proteins, fatty acids, and carotenoids from Chlorella, Spirulina, Aphanizomenon, and Chlamydomonas (Bleakley et al., 2017; Caporgno et al., 2018; Wang et al., 2021; Acquah et al., 2021; Pyne et al., 2022). Scenedesmus and Chlorella are used in bio-floc systems and fish feed supplements to enhance the growth of Pacific white shrimp and Nile tilapia (Silva et al., 2022).
Natural nitrogen fixators
Nitrogen is a crucial macronutrient for plant growth, development, and productivity (Huang et al., 2020). It is a vital component of chlorophyll, proteins, enzymes, and nucleic acids. However, plants cannot directly use atmospheric nitrogen gas, which makes up 78% of the air (Rilling et al., 2020). Microbes play a crucial role in fixing nitrogen through symbiotic or non-symbiotic relationships. Free-living bacteria such as Rhizobium, Azospirillium, Azobacter, and Bradyhizobium can colonize various plant rhizospheres, including those of Lupinus luteus and legumes (Yousuf et al., 2017; Saxena et al., 2019; Grover et al., 2021). These nitrogenase bacteria convert atmospheric nitrogen into ammonium, enabling plants to access and uptake nitrogen (Huang et al., 2020). The bioaugmentation or inoculation of plants with nitrogen-fixing bacteria reduces fertilization costs and is environmentally friendly. These bacteria are also valuable for improving the performance of grain crops and reducing our dependency on chemical nitrogen fertilization (Huang et al., 2020; Rilling et al., 2020).
Mineral solubilizers
The availability of soluble minerals in the soil is critical for enhancing crop growth and productivity. While fertilizers are a common source of minerals, the phosphorus they provide is often retained in the soil's solid phase. Phosphorus can form precipitates with aluminum, iron, calcium, and organic matter or be adsorbed onto the surface of clay particles, rendering it unavailable for plant uptake (de Abreu et al., 2017). However, rhizosphere-associated microbes such as Aspergillus, Penicillium, Trichoderma, and mycorrhizal fungi can solubilize soil-bound phosphorus (Mandal and Tiru, 2021). Soil inoculated with these microbes improves the availability of phosphorus for plant root uptake or absorption (Saxena et al., 2019; Mandal and Tiru, 2021). Phosphatesolubilizing bacteria (PSB), including Azotobacter chroococcum, Bacillus subtilis, Bacillus cereus, Bacillus megaterium, Arthrobacter ilicis, Escherichia coli, Pantoea agglomerans, Pseudomonas putida, Pseudomonas aeruginosa, Enterobacter aerogenes, Microbacterium laevaniformans, and Micrococcus luteus (de Abreu et al., 2017; Li et al., 2021), secrete organic acids such as citric, gluconic, lactic, propionic, and succinic acids. These organic acids help reduce soil pH and may act as chelators for divalent cations such as calcium, releasing bonded phosphorus iron for plant absorption (Saxena et al., 2019). For instance, Amri et al. (2022) found that using Pseudomonas fluorescens as a P-fertilizer on ryegrass increased root proliferation and plant biomass. This approach is promising and could serve as an alternative source of P-fertilizers to maintain high nutrient levels in crops such as maize, wheat, and rice (de Abreu et al., 2017; Li et al., 2021).
Production of nano-fertilizers
Physicochemical methods have been utilized to produce nanoparticles (NP) of various mineral elements for agricultural use (Ng et al., 2021). Nanoparticle-based fertilizers (NPF) offer an advantage over bulk or conventional fertilizers due to their easy absorption and transportation in plant vascular tissues. They can also be used as carriers for fertilizers and pesticides, which can enhance chemical efficiency and reduce operational costs (Rastogi et al., 2017, 2019; Ng et al., 2021; Noor-Hashim et al., 2021). The technique of using microbial enzymes (bacterial or fungal) in the production of NPs is referred to as “green synthesis of nanoparticles” (Zielonka et al., 2018). This method is particularly useful in the production of NPs for agricultural purposes. Silica nanoparticles (Si-NP) are an example of fertilizers that are successfully produced using Aspergillus parasiticus (Zielonka et al., 2018) or Trichoderma (review by Ramirez-Valdespino and Orrantia-Borunda, 2021). Additionally, the use of silicon has been reported to enhance the growth and productivity of rice, which is one of the main staple food crops (Zielonka et al., 2018; Ng et al., 2021; Noor-Hassim et al., 2021).
Plant growth-promoting microbes
In addition to soil mineral nutrients, research indicates that the use of beneficial microorganisms can promote plant health by increasing phytohormone levels in the rhizosphere (Teo et al., 2022). Plant growth-promoting microbes (PGPM) have a positive impact on crop growth and soil health without competing with other soil microflora. It is estimated that over 80% of soil bacteria have the potential to contribute to the exogenous pool of auxin (IAA) available for plant uptake (Afzal et al., 2019; Grover et al., 2020). The intensification of root hair formation, biomass, and root length was observed in the symbiotic association of PGPM with the crop-rhizosphere (Grover et al., 2021). PGPM also secrete gibberellins (GA) and cytokinins, which are other types of plant growth regulators (Teo et al., 2022). Gibberellin acts as a plant physiological regulator that promotes seed germination, primary root elongation, cell division, and elongation, all of which contribute to promoting plant growth (Backer et al., 2018). Cytokinin improves seedling vigor by promoting shoot growth (Grover et al., 2021; Jiao et al., 2021). PGPM, which includes bacteria such as Pseudomonas sp. and Bacillus, fungi, and actinobacteria, increases the solubilization of inaccessible forms of soil minerals into bioavailable forms (Arora et al., 2020; Zin and Badaluddin, 2020; Riaz et al., 2021). PGPM also improve crop performance by facilitating nitrogen fixation, mineral nutrient solubilization, chelation of minerals, ion-exchange reactions producing phytohormones and organic acid, and combating pathogen overproduction of biologically active metabolites (Zhang et al., 2017; Egamberdieva et al., 2019, 2021). The application of PGPM as biofertilizers is widely accepted as an eco-friendly approach to replace hazardous chemical inputs in agricultural systems to sustain intensification (Teo et al., 2022).
Biopesticides
The use of chemicals for controlling pests and diseases has been prevalent for decades, resulting in increased operating costs for farmers and the accumulation of residues in the environment (Arora et al., 2020; Verma et al., 2020; Lopes et al., 2021). To address this issue, the use of microbes with plant growth-promoting activity as natural biopesticides is gaining attention as a sustainable alternative (Verma et al., 2020). Microbes such as Bacillus, Pseudomonas, and Rhizobium secrete various compounds, including antibiotics, siderophores, and secondary metabolites, to eliminate plant pathogens (Backer et al., 2018; Kenawy et al., 2019; Verma et al., 2020; Riaz et al., 2021; Teo et al., 2022). In addition, siderophores-producing bacteria, such as Aeromonas, Azadirachta, Azotobacter, Bacillus, Burkholderia, Pseudomonas, Rhizobium, Streptomyces, and Serratia species, have been reported to activate plant systemic resistance genes (Romera et al., 2019; Sarwar et al., 2020; Sultana et al., 2021). Trichoderma has been shown to be an effective fungus in combating crop diseases caused by Bipolaris oryzae, Meloidogyne javanica and Rhizoctonia solani, which affect crops such as rice, tomatoes, and potatoes (Hariharan et al., 2022; Mukherjee et al., 2022). It produces active secondary metabolites such as anthraquinones, cyclopentenone, ergosterol derivatives, and N-heterocyclic compounds, which are effective against these crop pathogens (Meena et al., 2020; Scudeletti et al., 2021; Verma et al., 2021).
Pollution and heavy metal remediators
Soil health is crucial for crop yield production, but the intrusion of seawater into farmland due to rising seawater levels can harm crops. High salinity levels can negatively affect seed germination, root systems, and nutritional uptake, and cause imbalances that lower crop productivity (Arora et al., 2020; Litelien and Zeeb, 2020; Mwando et al., 2020). Stress-tolerant microbes show great potential for remediating polluted soil, improving soil health, and sustaining crop production (Egamberdieva et al., 2021). Trichoderma and Mycorrhiza, which colonize plant roots, protect plants from environmental stresses such as salinity, drought, and heavy metals (Siti-Atirah et al., 2020; Zhang et al., 2020; Scudeletti et al., 2021). Moreover, microbes like Azospirillum, Pseudomonas, and Bacillus exhibit versatile bio-sorbent characteristics under extreme soil pH, nutrient availability, and temperature (Lopes et al., 2021; Scudeletti et al., 2021). Rhizosphere-colonized microbes can also release siderophores (Sarwar et al., 2020; Egamberdieva et al., 2021). Siderophores help plants survive under extreme soil conditions such as high levels of mineral ions (Abdallah et al., 2021; Ramirez-Valdespino and Orrantia-Borunda, 2021; Sultana et al., 2022).
Soil-water binders
Drought poses a significant threat to crop production worldwide, and the relationship between plants, soil, and water is crucial to the seasonal water saturation of the soil. Soil with good drainage but low water-holding capacity can lead to severe water restrictions on plants. Mycorrhiza and Trichoderma have been reported to have a high soil-water-holding capacity, with their mycelium enhancing soil water repellence and being sufficient to maintain water-stable soil aggregate (Marimon-Junior et al., 2020; Zin and Badaluddin, 2020; Scudeletti et al., 2021). In addition, certain microbes such as Bacillus and Paenibacillus isolated from arid and desert soils produce lipopolysaccharides or lipoproteins that act as biosurfactants (BS) and bio-emulsifiers (BE), which are polyextremotolerant in various soils for up to 30 months after inoculation (Noura et al., 2018). Indeed, the application of BS or BE-producing microbes to improve the soil hydrological properties in arid regions to mitigate drought stress is an alternative approach to the conventional inoculation of rhizobacteria. Siderophores are also helpful in aiding plant survival under water scarcity conditions (Abdallah et al., 2021; Ramirez-Valdespino and Orrantia-Borunda, 2021; Sultana et al., 2022).
Conclusions
Microbes are one the important natural resources that should be utilized for sustainable food production. Microbes can be directly taken as human food and feed supplements for livestock. In agri-food production, microbes can be utilized as natural fertilizers, improving soil health, and biocides to protect crops from pathogens and diseases. Extensive utilization of microbes can reduce the dependence on synthetic fertilizers and pesticides for sustainable food production.