

Introduction
Type 1 diabetes (T1D) is one of the most common auto-immune diseases which develops due to genetic and environmental risk factors [1]. This disease is also known as insulin-dependent, juvenile or childhood-onset diabetes [2]. T1D is based on the immune-mediated loss of pancreatic β cells. This deprivation leads to insulin deficiency, which means that people suffering from this disease do not produce enough insulin and are characterized by the clinical need for it. Insulin-secreting cells can be destroyed by genetic and non-genetic factors [1]. Clinical symptoms occur when β cell deficiency exceeds 40% of their overall amount. This is combined with detectable decrease in concentration a marker of pancreatic insulin production, plasma C-peptide [3]. What is more, most of T1D patients exhibit a presence of disease-associated serum autoantibodies [1]. Nowadays, T1D diagnosis is partially based on the detection of islet cells autoantibodies. The most often detected are the following antibodies: 1) insulin autoantibodies (IAA), 2) anti-glutamic acid decarboxylase (GAD) autoantibodies, 3) protein tyrosine phosphatase-related islet antigen 2 (IA-2) autoantibodies (also known as ICA512 autoantibodies) [4] and 4) zinc transporter 8 (ZnT8) auto-antibodies [3]. It has been already proven that specific human leukocyte antigen (HLA) haplotypes are associated with islet primary autoantibodies, which are observed at seroconversion. For example, HLA-DR4-DQ8 is associated with IAA and HLA-DR3-DQ2 is associated with GAD [5]. These circulating autoantibodies are characteristic of T1D type 1A. A much less common type (roughly 10% of cases) of T1D is type 1B (alias idiopathic T1D). Patients suffering from this type of diabetes are autoantibody-negative, which might suggest that they suffer from 1) a rare (about 6% of T1D cases) monogenic form of diabetes or 2) a lack of measurable autoantibodies responses to common antigens [6, 7].
T1D occurs in genetically susceptible patients that were subjected to one or more triggering environmental factors which induced an autoimmune process leading to destruction of β cells in islets of Langerhans [8-11]. The preclinical period of antibody production and islet destruction can last several months or even years and it is crucial in terms of medical intervention focused on slowing down or stopping this process [12-14]. Early identification of people with high genetic predisposition to T1D extends the time for prevention or treatment and helps to decelerate the disease [15-17]. Maybe in the future we will be able to stop the changes in the immune system and the development of autoantibodies that take part in the destruction process of insulin-producing cells [1].
Polymorphisms of multiple genes are reported to modify the risk and course of T1D [17-20]. Among them, numerous variants of genes from the HLA region are factors playing a very important role in T1D. They can affect the presence of autoreactive antigen T lymphocytes during a diabetic inflammatory process [21-24]. Coexistence of various haplotypes affecting autoimmune response and balance between them might determine the onset time, the process and dynamics of the disease or its clinical form [20, 21, 25].
Identification of factors implicated in diabetes and mechanisms of interaction between those factors should lead to a better understanding of the disease pathogenesis and help in the development of T1D prevention strategies.
Genetic studies in diabetes
T1D is a member of the multifactorial disease group [22]. One of its most important factors are variants of HLA region. First data suggesting the significance of associations in the HLA region were published in the 1970s [23, 24]. Many studies have been performed since then identifying associations not only in genes constituting the HLA region but also outside this region [25]. The single nucleotide polymorphism (SNPs), which are associated with islet autoimmunity and high risk of T1D, are placed in such non-HLA genes as PTPN22 (protein tyrosine phosphatase non-receptor type 22), UBASH3A (ubiquitin associated and SH3 domain containing A), ERBB3 (Erb-B2 receptor tyrosine kinase 3), CLEC16A (C-type lectin domain containing 16A), IL27 (interleukin 27), CTRB (chymotrypsinogen B1), C14orf64 [also known as LINC01550 (long intergenic non-protein coding RNA 1550)], GSDM (gasdermin A) and HORMAD2 [HORMA (Hop1, Rev7 and MAD2) domain containing 2] [4]. Three technical approaches are used for detection of risk loci in T1D: linkage studies, association studies and genome-wide association studies (GWAS).
High risk loci identified by these three approaches explain almost 80% of the heritability of T1D. To get a better understanding of T1D, further genetic studies are required to explain how genetic variants change β-cell response to environmental factors or inflammatory mediators. This can become possible with the use of new technologies like next generation sequencing (NGS).
Linkage studies
Linkage studies are capable of identifying genomic regions which are shared more frequently between affected relatives, and they generally examine sibling pairs [26]. This approach is mostly used when effect sizes of the risk factors are relatively large (compared to GWAS) while analyzed markers are present in a genome in small density. And for this reason it is worth analyzing those markers further because they may play a much more important role in T1D. The HLA region (6p21) is a good example and an evidence of linkage with T1D and therefore it proves the importance of additional research [27, 28].
Association studies
Association studies allow us to detect alleles that are rather coon but they have a modest effect on a disease risk [26]. In the simplified form, association studies compare frequency of an allele/a genotype of a specific variant between disease cases and a control group [29]. They are generally focused on candidate genes [30]. T1D association studies identified 4 non-HLA genes with established risk loci [HLA, INS (insulin), CTLA4 (cytotoxic T-lymphocyte antigen 4), PTPN22] [4].
Genome-wide association studies
Emergence of GWAS revolutionized genetic studies and genome examination in search of risk factors. GWAS lets us survey a genome in search of causal variants i.e. variants that contribute to a disease but may not be a sufficient reason for this disease in isolation [31]. Analyses performed by The Welcome Trust Case Control Consortium (WTCCC) [32] and Type 1 Diabetes Genetics Consortium [33, 34] have revealed more than 50 genetic variants associated with T1D development [19]. These studies, next to well-established non-HLA loci, have reported many new associations with T1D, among them: GLIS3 (GLIS family zinc finger 3), LMO7 (LIM Domain Only 7), FUT2 (fuco-syltransferase 2) [35-37]. Additionally, genome-wide genotyping approach of nonsynonymous SNPs (nsSNP), a direct precursor of GWAS, uncovered the sixth gene ever associated with T1D – IFIH1 (interferon induced with helicase C domain 1) [38, 39].
Candidate genes for type 1 diabetes
Susceptibility to T1D is strongly associated with a genetic background. GWAS contributed to the discovery of a number of T1D susceptibility genes. Before GWAS only five loci were determined as associated with T1D. They were (in that order): HLA region, INS, CTLA4, PTPN22 and IL2RA. GWAS meta-analyses have identified more than 50 T1D susceptibility loci (Fig. 1). Some of these genes can act as modulators of an immune-related process through both antigen presentation and other modifications of the immune function. A number of candidate genes involved in T1D are known as modulators of β-cell apoptosis, viral infection or islet inflammation. Some of the main factors causing changes in susceptibility to T1D are SNPs and variations in gene expression (genetic variants in regulatory elements of genes can result in alteration of transcription and then gene expression). In addition, most of the identified genes are expressed both in immune competent cells and in pancreatic β-cells, which suggests a genetically modulated dialogue between these two components of T1D. Below we present recently examined genes associated with T1D development according to GWAS study with the statistical threshold for genome-wide significance, P-values < 5 × 10–8 (P-values calculated using Fisher’s combined p-value method implemented in Haploview) [25]. In Table 1 we characterised genes which have been indicated as ones associated with T1D in studies performed so far. Their SNPs positions on chromosomes are illustrated in Figure 1.
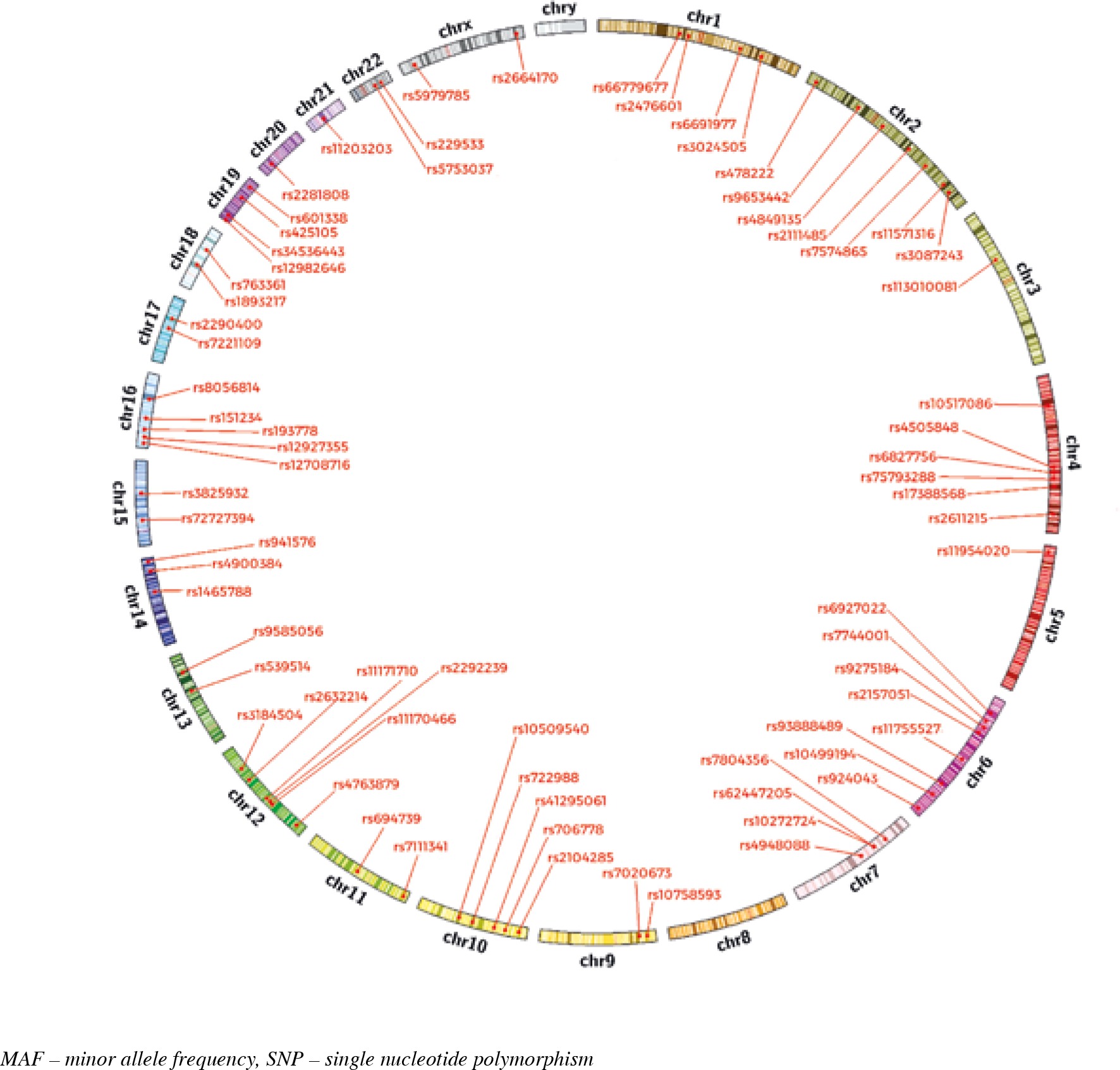
Table 1
Type 1 diabetes associated loci according to ImmunoBas [91] and the latest studies
| Region | Gene | SNP | HGVS nomenclature [92] | MAF | Coding protein | Role | Ref. |
|---|---|---|---|---|---|---|---|
| 1p13.2 | PTPN22 PHTF1 | rs2476601 rs6679677 | NC_000001.11:g.113834946A>G NC_000001.10:g.114303808C>A | A = 2.74% A = 2.58% | Protein tyrosine phosphatase, non-receptor type 22 Putative homeodomain transcription factor 1 | Lymphoid protein tyrosine phosphate is a key molecule regulating TCR signaling and the SNP in these gene could leads to higher T-cell activation PHTF1 plays role in transcription regulation | [91] |
| 1q32.1 | CAMSAP2 | rs6691977 | NC_000001.10:g.200814959T>C | C = 42.45% | Calmodulin regulated spectrin associated protein family member 2 | Not applicable | [93] |
| 1q32.1 | IL10 | rs3024505 | NC_000001.10:g.206939904G>A | A = 8.63% | Interleukin 10 | IL-10 is an anti-inflammatory cytokine | [91] |
| 2q24.2 | IFIH1 | rs2111485 | NC_000002.11:g.163110536A>G | G = 33.93% | Interferon induced with helicase C domain 1 | The protein acts as cytoplasmic sensor of viral nucleic acids and plays role in sensing viral infection and in the activation of cascade of antiviral responses by induces of type I interferons and proinflammatory cytokines. SNP in these gene may lead to increase viral infection and innate immune response | [93] |
| 2q32.3 | STAT4 | rs7574865 | NC_000002.11:g.191964633T>G | T = 25.54% | Signal transducer and activator of transcription 4 | STAT4 plays role as a signal transduction and activation of transcription. SNP in these gene is associated with several autoimmune disorders but there is no know exactly role of these mutation in developing of disease | [65] |
| 2q33.2 | CTLA4 | rs11571316 rs3087243 | NC_000003.11:g.102220660A>G NC_000002.11:g.204738919G>A | A = 47.1% A = 36.9% | Cytotoxic T-lymphocyte associated protein 4 | CTLA4 is an protein receptor and downregulates immune reaction | [25, 91] |
| 2q13 | ACOXL | rs4849135 | NC_000002.11:g.111615079T>G | T = 30.7% | acyl-CoA oxidase like | Not applicable | [93] |
| 2p23.3 | EFR3B | rs478222 | NC_000002.11:g.25301755A>T | T = 36.74% | EFR3 homolog B | The protein is involved in signaling processes | [25] |
| 2q11.2 | AFF3 | rs9653442 | NC_000002.11:g.100825367C>T | T = 43.11% | AF4/FMR2 family member 3 | Transcriptional activator, involved in lymphoid development and oncogenesis | [94] |
| 3p21.31 | CCR5 | rs113010081 | NC_000003.11:g.46457412T>C | C = 2.84% | C-C chemokine receptor 5 | The CCR5 affects the immune cell function, mutation in this gene influences the function of immune cells response | [93] |
| 4q32.3 | AC080079.1 or NOL8P1 | rs2611215 | NC_000004.11:g.166574267A>G | A = 18.17% | Tripartite motif-containing 38 (TRIM38) pseudogene or nucleolar protein 8 pseudogene 1 | Not applicable | [93] |
| 4q27 | ADAD1 IL21 IL2 | rs6827756 rs4505848 rs17388568 | NC_000004.12:g.122263256T>A NC_000004.12:g.122211337A>G NC_000004.12:g.122408207G>A | T = 40.83% G = 29.63% A = 14.06% | Adenosine deaminase domain containing 1 Interleukin 21 Interleukin 2 | ADA1 plays role in spermatogenesis and binds to RNA IL-21 binds to receptors on B and T lymphocyte and NK cells IL-2 is the most important growth factor for developing T lymphocytes | [32, 94] |
| 4p15.2 | LINC02357 | rs10517086 | NC_000004.11:g.26085511G>A | A = 18.59% | Long intergenic non-protein coding RNA 2357 | Not applicable | [91] |
| 5p13.2 | IL7R | rs11954020 | NC_000005.9:g.35883251C>G | G = 39.92% | Interleukin 7 receptor | This receptor plays a key role in developing lymphocytes | [93] |
| 6q22.32 | CENPW | rs9388489 | NC_000006.11:g.126698719A>T | A = 44.81% | Centromere protein W | One of the components of the CENPA-NAC complex which plays key role in assembly of kinetochore protein | [95] |
| 6q15 | BACH2 | rs11755527 | NC_000006.11:g.90958231C>G | G = 36.66% | BTB domain and CNC homolog 2 | Plays role in coordinating transcription activation and repression. Induces apoptosis in response to oxidative stress | [96] |
| 6q27 | AL596442.1 or AL049612.1 | rs924043 | NC_000006.11:g.170379025T>C | T = 41.47% | Novel transcript | Not applicable | [25] |
| 6q23.3 | TNFAIP3 | rs6920220 rs10499194 | NC_000006.11:g.138006504G>A NC_000006.11:g.138002637C>T | A = 9.44% T = 19.15% | TNF-α induced protein 3 | Involved in immune and inflammatory responses signaled by cytokine such as TNF-α or by pathogens | [73] |
| 6p21.3 | HLA class II | rs6927022 rs2157051 rs9275184 rs7744001 | NC_000006.11:g.32612397A>G NC_000006.11:g.32658624G>A NC_000006.11:g.32654714T>C NC_000006.11:g.32626086G>C | G = 35.76% G = 39.08% C = 8.71% A = 38.92% | Major histocompatibility complex | Binds peptide derived from antigens and presents them on the cell surface for recognition by the CD4 T-cells | [97] |
| 7p15.2 | SKAP2 | rs7804356 | NC_000007.13:g.26891665T>C | C = 17.83% | Src kinase associated phosphoprotein 2 | Not applicable | [91] |
| 7p12.1 | COBL | rs4948088 | NC_000007.13:g.51027194A>C | A = 3.99% | Cordon-Bleu WH2 repeat protein | Involved in the reorganization of the actin cytoskeleton and plays key role in morphogenetic processes of the central nervous system | [98] |
| 7P12.2 | IKZF1 | rs10272724 rs62447205 | NC_000007.13:g.50477213T>C NC_000007.13:g.50465830A>G | C = 21.33% G = 22.62% | IKAROS family zinc finger 1 | This protein binds to DNA and plays role as a transcriptional regulator of hematopoietic cell differentiation | [93, 99] |
| 9p24.2 | GLIS3 | rs7020673 rs10758593 | NC_000009.11:g.4291747C>G NC_000009.11:g.4292083G>A | C = 35.82% A = 47.92% | GLIS family zinc finger 3 | Plays a role in generation of pancreatic beta cells and in insulin gene expression | [100] |
| 10p11.22 | NRP1 | rs722988 | NC_000010.10:g.33426147T>C | T = 44.95% | Neuropilin 1 | Involved in axonal growth and guidance and in physiological angiogenesis | [101] |
| 10p15.1 | IL2RA | rs41295061 rs2104285 rs706778 | NC_000010.11:g.6072697C>A NC_000010.11:g.5868542G>A NC_000010.11:g.6056986C>T | A = 2.8% T = 32.61% C = 49.92% | Interleukin 2 receptor subunit α | The receptor plays role in the immune tolerance by controlling regulatory T cells activity | [60, 102] |
| 10q23.31 | RNLS | rs10509540 | NC_000010.11:g.88263276T>C | C = 24.74% | Renalase FAD dependent anime oxidase | This enzyme degrades catecholamines such as adrenaline in the blood circulation and may regulates blood pressure | [103] |
| 11p15.5 | INS | rs7111341 | NC_000011.10:g.2191936C>T | T = 23.86% | Insulin | Decreases blood glucose concentration | [104] |
| 11q13.1 | BAD | rs694739 | NC_000011.10:g.64329761A>G | G = 21.17% | BCL2 associated agonist of cell death | Promotes cell death by initiating apoptosis | [101] |
| 12q13.13 | ITGB7 | rs11170466 | NC_000012.11:g.53585859C>T | T = 6.43% | Integrin subunit β7 | Adhesion molecule, mediates lymphocyte migration and homing to gut-associated lymphoid tissue (GALT). Interacts with fibronectin, an extracellular matrix component | [101] |
| 12q13.2 | ERBB3 DGKA | rs2292239 rs11171710 | NC_000012.11:g.56482180T>G NC_000012.11:g.56368078G>A | T = 6.43% A = 35.18% | Erb-B2 receptor tyrosine kinase 3 Diacylglycerol kinase α | ERBB3 plays role as cell surface receptor for molecules involved in developing of nervous system and embryogenesis DGKA converts a second messenger diacylglycerol into phosphatidate | [101, 105] |
| 12q21.2 | ZDHHC17 | rs2632214 | NC_000012.11:g.77032629T>C | C = 25.66$ | Zinc finger DHHC-type containing 17 | Probably involved in the sorting of critical proteins involved in the initiating endocytosis at the plasma membrane. | [36] |
| 12q24.13 | SH2B3 | rs3184504 | NC_000012.11:g.111884608T>C | T = 14.74% | SH2B adaptor protein 3 | This protein functions as a regulator in signaling pathways relating to inflammation, hematopoiesis and cell migration | [104] |
| 12p13.31 | CD69 | rs4763879 | NC_000012.11:g.9910164G>A | A = 31.29% | CD69 molecule | Involved in lymphocyte proliferation and functions as a signal-transmitting receptor in lymphocytes | [91] |
| 13q22.2 | LMO7 | rs539514 | NC_000013.10:g.76326282A>C | A = 26.78% | LIM domain 7 | Probably LMO7 is involved in protein-protein interaction | [25] |
| 13q32.3 | GPR183 | rs9585056 | NC_000013.10:g.100081766C>T | C = 28.23% | G protein-coupled receptor 183 | The protein is expressed in lymphocytes and acts as chemotactic receptor for B-cells, T-cells, splenic dendritic cells | [93] |
| 14q32.2 | AL163932.1 or LINC01550 | rs4900384 | NC_000014.9:g.98032614A>G | G = 49.76% | Novel transcript or long intergenic non-protein coding RNA 1550 | Not applicable | [91] |
| 14q24.1 | ZFP36L1 or MAGOH3P | rs1465788 | NC_000014.9:g.68796882T>C | T = 28.81% | ZFP36 ring finger protein like 1 or mago homolog 3, pseudogene | Not applicable | [91] |
| 14q32.2 | DLK1 | rs941576 | NC_000014.9:g.100839708A>G | G = 37.84% | Delta like non-canonical notch ligand 1 | May have a role in an neuroendocrine differentiation. | [106] |
| 15q25.1 | CTSH | rs3825932 | NC_000015.9:g.79235446T>C | C = 36.8% | Cathepsin H | It is a lysosomal cysteine proteinase important for the overall degradation of proteins in lysosomes | [91] |
| 15q14 | RASGPR1 | rs72727394 | NC_000015.9:g.38847022C>T | T = 13.78% | RAS guanyl releasing protein 1 | It regulates T-cells and B-cells development, homeostasis and differentiation | [93] |
| 16p11.2 | IL27 | rs151234 | NC_000016.9:g.28505660G>C | C = 13.20% | Interleukin 27 | It has pro- and anti-inflam- matory properties that, regulate T-helper cell development, suppress T-cell proliferation and has diverse influence on innate immune cells | [93] |
| 16p13.13 | CLEC16A DEXI | rs12708716 rs12927355 rs193778 | NC_000016.9:g.11179873A>G NC_000016.9:g.11194771C>T NC_000016.9:g.11351211A>G | G = 35.08% T = 27.16% G = 14.88% | C-type lectin domain containing 16A Dexamethasone-induced protein | Regulator of mitophagy, a selective form of autophagy necessary for mitochondrial quality control DEXI functions as an anti-inflammatory and immunosuppressant | [91, 93] |
| 16q23.1 | BCAR1 | rs8056814 | NC_000016.9:g.75252327G>A | A = 18.09% | Breast cancer anti-estrogen resistance 1 | Plays a role in coordinating for tyrosine kinase-based signaling related to cell adhesion. Involved in induction of cell migration | [93] |
| 17q21.2 | SMARCE1 | rs7221109 | NC_000017.11:g.40614034T>C | T = 29.59% | SWI/SNF related, matrix associated, actin dependent regulator of chro- matin, subfamily e, member 1 or TEC | Not applicable | [91, 107] |
| 17q12 | ORMDL3 | rs2290400 | NC_000017.10:g.38066240T>C | C = 42.05% | ORMDL sphingolipid biosynthesis regulator 3 | Negative regulator for sphingolipid synthesis and could plays role in regulate endoplasmic reticulum-mediated Ca2+ signaling | [91] |
| 18q22.2 | CD226 | rs763361 | NC_000018.9:g.67531642T>C | C = 46.94% | CD226 molecule | Involved in intracellular adhesion, lymphocyte signaling, cytotoxicity, stimulates T-cell proliferation and cytokine production | [91] |
| 18p11.21 | PTPN2 | rs1893217 | NC_000018.9:g.12809340A>G | G = 11.96% | Protein tyrosine phosphatase, non-receptor type 2 | Negatively regulates signaling pathways and biological processes such as hematopoiesis, inflammatory reactions, cell proliferation and glucose homeostasis | [91] |
| 19q13.33 | FUT2 | rs601338 | NC_000019.9:g.49206674G>A | A = 32.17% | Fucosyltransferase 2 | The protein is Golgi stack membrane protein that plays role in the creation of a precursor of antigen H. It mediates the transfer of fucose to the terminal galactose on glycan chains of cell surface glycoproteins and glycolipids | [37] |
| 19q13.32 | PRKD2 | rs425105 | NC_000019.9:g.47208481T>C | C = 14.26% | Protein kinase D2 | Not applicable | [91] |
| 19p13.2 | TYK2 | rs34536443 | NC_000019.9:g.10463118G>C | C = 1.02% | Tyrosine kinase 2 | Involved in intracellular signal transduction, may plays role in anti-viral immunity | [108] |
| 19p13.3 | CDC34 MADCAM1 | rs12982646 | NC_000019.9:g.499978G>A | A = 15.73% | Ubiquitin conjugating enzyme E2 R1 Mucosal vascular addressin cell adhesion molecule 1 | CDC34 catalyzes the covalent attachment of ubiquitin to other proteins, is involved in degradation of cell cycle G1 regulators MADCAM1 is a cell adhesion leukocyte receptor expressed by mucosal venules and helps lymphocytes traffic into mucosal tissues | [101] |
| 20p13 | SIRPG | rs2281808 | NC_000020.10:g.1610551T>A | T = 21.29% | Signal regulatory protein gamma | Not applicable | [91] |
| 21q22.3 | UBASH3A | rs11203203 | NC_000021.9:g.42416077G>A | A = 19.79% | Ubiquitin associated and SH3 domain containing A | Promotes accumulation of activated receptors, like T-cell receptors on the cell surface | [91] |
| 22q12.2 | AC002378.1 | rs5753037 | NC_000022.10:g.30581722C>T | T = 27.88% | novel transcript | Not applicable | [91] |
| 22q12.3 | C1QTNF6 RAC2 | rs229533 | NC_000022.10:g.37587111A>C | A = 44.59% | C1q and TNF related 6 Ras-related C3 botulinum toxin substrate 2 | Rac2 is a signaling G protein, regulates diverse cellular processes, controls cell growth and activation of protein kinases | [93] |
| Xp22.2 | TLR7/8 | rs5979785 | NC_000023.10:g.12971524C>T | C = 42.62% | Toll like receptor 7/8 | TLR family plays a role in pathogen recognition and activation of immune responses | [71] |
| Xq28 | GAB3 | rs2664170 | NC_000023.10:g.153945602G>A | G = 35.26% | GRB2 associated binding protein 3 | Not applicable | [91] |
The HLA region involved in progress of type 1 diabetes
The first connection between genetics and T1D that came into view was the HLA region located on chromosome 6p21. It explains around 50% of the overall heritability of the disease, therefore it is the strongest determinant for T1D. Genes from that region play multiple roles during immune response and they are the first checkpoints in its activation. Genetic polymorphisms encoding different amino acid residues in the peptide-binding pockets of HLA molecules are the main connection between HLA molecules and T1D. Moreover, the binding repertoire and affinity of peptides can be presented on T-cells [6, 40]. There are 2 classes of HLA genes: 1) class I – HLA-A, HLA-B and HLA-C, 2) class II – HLA-DR, HLA-DQ and HLA DP (HLA-DRB1, HLA-DQA1, HLA-DQB1, HLA-DPA1, HLA-DPB1) [5].
The strongest association with T1D is located within the HLA class II region genes that encode highly polymorphic β-chains (HLA-DRB1, -DQA1 and -DQB1) [41]. There are two main high-risk haplotypes “DR4-DQ8” (DR4-DQA1*03:01-DQB1*03:02) and “DR3-DQ2” (DRB1*03:01-DQA1*05:01-DQB1*02:01). Around 90% of T1D patients carry DR4-DQ8 or DR3-DQ2 and roughly 30% patients carry the combination of both of those haplotypes (DR4-DQ8/DR3-DQ2). This group confers the highest risk of T1D development (OR = 16) [6, 42].
HLA class I alleles are also responsible for T1D but less strongly. The most associated genes from that group are HLA-A and HLA-B [43]. The presence of the HLA-B*39 allele has been shown as a significant risk factor for the disease. Moreover, it is connected with T1D diagnosis at young age [44]. In addition, HLA-A*02 also increases the likelihood of T1D development and is one of the most frequent class I alleles, with frequency of > 60% in T1D patients [45].
The non-HLA genes associated with type 1 diabetes
The insulin gene (INS) encoded on chromosome 11p15 was the second locus linked to T1D. One of the main reasons for insulin gene association with the disease is the existence of the VNTR (variable number tandem repeat) polymorphisms. Polymorphisms of this region have been divided into three classes according to the amount of nucleotide repeats: class I (26-63 repeats), class II (around 85 repeats) and class III (120-170 repeats) [46]. All these polymorphisms are responsible for the amount of insulin mRNA in the thymus. Class I VNTRs (the shortest) present the highest risk of T1D while class III VNTRs are believed to protect against T1D [47-49].
The next recognized locus was the cytotoxic T-lymphocyte antigen 4 (CTLA4 gene) encoded on chromosome 2q33.2, which belongs to the immunoglobulin superfamily and binds to the B7 molecule on antigen-presenting cells. A study showed that the A49G polymorphism [A to G transition at position 49 of the first exon NC_000002.12:g.203867991A>G (rs231775)] is the most associated with T1D (from all CTLA4 SNPs) [50, 51]. There are two more CTLA4 gene variants discovered: 1) C to T transition at position – 319 of the promoter region and 2) (AT)n dinucleotide repeat polymorphism at position 642 of the 3’ UTR. Both of those polymorphic markers did not show as strong association with T1D as NC_000002.12:g.203867991A>G in the meta-analysis [52].
PTPN22 is another gene involved in T1D, strongly correlated with its development. It is located on chromosome 1p13. This gene encodes tyrosine-protein phosphatase non-receptor type 22 (also known as lymphoid tyrosine phosphatase – LYP), a negative regulator of T-cells receptor signaling. A SNP at position 1858 [NC_000001.11:g.113834946A>G (rs2476601)] encoding Arg620Trp is associated with the induction of islet autoantibodies and diabetes development [53, 54]. Its association with T1D was confirmed after observing its role in disruption of T-cell deactivation mechanism that leads to expansion of autoreactive T-cells [35].
The interleukin 2 receptor α (IL2RA; also known as CD25) gene on chromosome 10p15 is the fifth discovered T1D locus [55]. Protein encoded by this gene forms one subunit of the receptor for interleukin 2. This receptor takes part in controlling TREGs (regulatory T-cells) activity, whereas TREGs are responsible for the activation and expansion of autoreactive T-cells [56]. First study investigating IL2RA locus association with T1D was performed using a tag single nucleotide polymorphism approach [55]. Together with GWAS five SNPs were predominantly determined and presented a significant T1D association: NC_000010.11:g.6056986C>T (rs706778), NC_000010.11:g.6059750T>C (rs3118470) [57], NC_000010.11:g.6072697C>A (rs41295061), NC_000010.11:g.6080046T>A (rs11594656) [58] and NC_000010.11:g.6057082T>C (rs2104286) [59]. All those SNPs are considered T1D risk factors, though the strongest association was distinguishable for NC_000010.11:g.6080046T>A, NC_000010.11 :g.6057082T>C and NC_000010.11:g.6072697C>A, all of which were proved to be separately linked to the circulating concentration of IL2RA soluble form [58, 60].
The sixth identified gene and the first in the GWAS era was IFIHI (early type I interferon β-responsive gene) on chromosome 2q24.3. It was one of four genes in 2q.24 region with revealed association but it turned out to be the most noteworthy because of the protein it encodes. IFIH1 encodes interferon-induced helicase C domain-containing protein 1 (also called MDA5 – mela-noma differentiation-associated protein 5), which is an intracellular pathogen receptor. This discovery posed an interesting link between viral infections and T1D development and made IFIH1a particularly functional candidate for T1D disease [9, 38, 61]. The first association of IFIH1 was discovered through an interim analysis of a genome-wide nsSNP (direct precursor of GWAS) – NC_000002.12:g.162267541C>T (rs1990760) that leads to the A946T substitution [38]. Further research into this genetic variation led to the discovery postulating that A®T substitution in IFIH1 protein takes part in limiting viral infection (higher expression of type 1 interferons) but at the same time it promotes the risk of autoimmunity [62]. In the later years four rare variants of IFIH1 were discovered – NC_000002.12:g.162268127T>C (rs35667974), NC_000002.12:g.162279995C>G (rs35337543), NC_000002.12:g.162268086C>T (rs35732034) and NC_000002.12:g.162277580C>G (rs35744605) – which were proven to lower the risk of T1D independently of each other [63]. Moreover, another group of researchers showed three more SNPs with a distinct T1D association: one in the coding region [NC_000002.12:g.162272314T>C (rs3747517)] and two in the 3’ intergenic region of IFIH1 [NC_000002.12:g.162243749G>A (rs13422767) and NC_000002.12:g.162254026A>G (rs2111485)] [64].
The signal transducer and activator of transcription (STAT4) is a central mediator involved in generating inflammation during protective immune responses and immune-mediated disease. This gene is located on chromosome 2q32.2-q32.3 and studies showed that NC_000002.12:g.191099907T>G (rs7574865) polymorphism is associated with diabetes risk, especially T1D [65]. This polymorphism might affect STAT4 production or phosphorylation processes but this question has yet to be investigated.
The ubiquitin associated and SH3 domain containing A (UBASH3A) is a gene located on chromosome 21q22.3 and it encodes one of two family members belonging to the T-cell ubiquitin ligand (TULA) family that can negatively regulate T-cell signaling. Genetic variants of this gene have been proven to be involved in T1D. Studies demonstrated that SNPs, including NC_000021.9:g.42416077G>A (rs11203203) and NC_000021.9:g.42415901T>C (rs80054410), display significant associations with T1D [66]. These two risk variants in UBASH3A regulate expression of UBASH3A and IL2 in human primary CD4+ T cells.
The SH2B adapter protein 3 (SH2B3) gene located on locus 12q24 encodes a protein involved in signaling activities through growth factors and cytokine receptors. The nonsynonymous SNP NC_000012.12:g.111446804T>G (rs3184504) is a gene variant the most associated with T1D development (among all SH2B gene variants) [67]. This polymorphism can lead to novel SH2B3 isoforms that affect β-cell inflammation and death in vivo or modulate the immune system, however it is still unclear. Nonetheless, new insights into SH2B adapter protein 3’s function were shown recently. It was proved that it is responsible for controlling the homeostasis in adipose tissue. What is more it reduces the risk of diabetes through regulation of IL-15-dependent adipose G1-ILC [68].
The TLR7 and TLR8 genes (toll-like receptor 7 and 8) are located on one region of X chromosome (Xp22.2). They encode type I transmembrane proteins belonging to Toll-like receptor family and play a fundamental role in pathogen recognition and activation of innate immunity. NC_000023.11:g.12953405C>T (rs5979785) SNP was located 30 kb centromeric of those two genes and it presents some level of association with T1D [69-71]. It is highly probable that this SNP may modify the expression of both TLR7 and TLR8, which are functional T1D candidate genes based on their role as pathogen recognition receptors. Moreover, TLR7 overexpression has been associated with murine autoimmune disease directly [72].
The TNFAIP3 (Tumor necrosis factor α-induced protein 3) gene located on chromosome 6q23 with its two SNPs [NC_000006.12:g.137685367G>A (rs6920220) and NC_000006.12:g.137681500C>T (rs10499194)] is also associated with T1D disease. Both of those SNPs were shown to be linked to T1D independently of each other [73]. TNFAIP3 encodes a ubiquitin-editing enzyme (A20) which inhibits NF-κB activation and TNF-mediated apoptosis and acts as an anti-apoptotic protein in specific cell types [74].
The Huntingtin-interacting protein gene (HIP14/ZDHHC17) expressed in β-cells is required for β-cells survival and is a target of proinflammatory cytokines which contribute to β-cells destruction in T1D. HIP14 is a palmitoyltransferase specific for a subset of neuronal proteins and is important for intracellular trafficking and exocytosis in neurons [75]. A study showed that decreased level of HIP14 in β-cells led to increased cell apoptosis and disease development. Knockdown of HIP14 causes a decrease in release of insulin and therefore it suggests that HIP14 is important for β-cell insulin release [36].
The FUT2 gene located on chromosome 19q13.33 encodes a protein α-1,2-fucosyltransferase which is responsible for the creation of H antigen, that is required for the final step in the soluble A and B antigen synthesis. Studies demonstrated that a single nucleotide polymorphism in this gene [NC_000019.10:g.48703417G>A (rs60133)] encodes the “non-secretor” variant which is associated with T1D [37].
The LMO7 (LIM Domain Only 7) gene located on locus 13q22 encodes a protein containing a calponin homo-logy (CH) domain, a PDZ domain [PSD-95 (post-synaptic density protein 95); Dlg1 (Drosophila disc large tumor suppressor)]; ZO-1 (zona occludens 1)] and a LIM domain [Lin11 (Protein lin-11); Isl-1 (Insulin gene enhancer binding protein); Mec-3 (Mechanosensory protein 3)], and it may be involved in protein-protein interactions. This gene is expressed in pancreatic islets and thus it is a candidate for a gene associated with T1D. Studies showed that the SNP NC_000013.11:g.75752146A>T (rs539514) is involved in the development of disease [25].
The EFR3B (protein EFR3 homology B) gene is located on chromosome 2p23. Its protein is a component of a complex required to localize phosphatidylinositol 4-kinase (PIL4) on the plasma membrane and thus EFR3B presumably plays a role in membrane-anchoring [76]. EFR3B, like LMO7, is a newly discovered gene modulating susceptibility to T1D. The factor involved in the progress of T1D is SNP NC_000002.12:g.25078886A>T (rs478222) [25].
The Kruppel-like zinc finger protein Gli-similar (GLIS3) is a candidate gene involved in T1D. GLIS3 is located on 9p24.2 locus and encodes a nuclear protein with five C2H2-type zinc finger domains. This protein functions as a transcription activator and repressor and is also entangled in development of pancreatic β-cells. Mutations in this transcription factor cause neonatal diabetes and hypothyroidism [77]. GWAS studies showed that SNPs in this gene are associated both with T1D and T2D. What is more, knockdown of GLIS3 leads to increased apoptosis of islet cells both in basal condition and following an exposure to proinflammatory cytokines.
Non-coding RNAs in type 1 diabetes
A considerable part of the genome is transcribed but only a little portion of DNA encodes proteins. The non-protein-coding part of the genome is transcribed to create a wide spectrum of non-coding RNAs including tRNAs (transfer RNA), rRNAs (ribosomal RNA), micro RNAs (miRNAs) and long non-coding RNAs (lncRNAs) [78].
Long non-coding RNA
Long non-coding RNAs are 200 nucleotides or longer. They are capped, polyadenylated and spliced, in the same way as protein coding transcripts. The studies have shown that lncRNAs play a significant role in a variety of biological processes such as transcription, splicing, translation and apoptosis [79]. A number of mammalian lncRNAs are expressed in a cell-type specific way. The role of these parts of the genome as transcription factors suggests that lncRNAs could be mediators of lineage-specific differentiation or specialized cellular functions. Mutations in lncRNAs could be a potential factor in disease development and cell-specific regulatory lncRNAs might provide therapeutic targets. The latest studies have demonstrated that human islet lncRNAs are highly cell-type specific and they mediate biological processes in islet cells such as β-cell differentiation, insulin biosynthesis and insulin secretion [80].
So far, GWAS studies have identified a number of disease-associated single nucleotide polymorphisms not limited to protein-coding genes. The study from 2015 has shown that lncRNAs play a role in the autoimmune process and autoimmune diseases [81]. The analysis of non-coding portion of the genome is important to identify novel biomarkers of disease development. According to transcriptome profiling studies of pancreatic islets, there are 1000 islet-specific lncRNAs, which are common for human and mouse islets. SNPs within lncRNA regions might modify their secondary structure and change regulatory functions, which could lead to an increased risk of developing diabetes. It has been presented that HI-LNC25 lncRNA [also known as LINC01370 (long intergenic non-protein coding RNA 1370)] is a regulator of GLIS3 mRNA [80]. Knockdown of HI-LNC25 caused reduced mRNA levels of GLIS3, a gene encoding an islet transcription factor. Genetic variations of this gene can lead to T2D development. Another study indicated a link between lncRNAs and phenotype-associated loci within type 1 diabetes candidate loci. The SNP of NONHSAG044354 lncRNA located within BACH2 gene [NC_000006.12:g.90247744C>T (rs3757247)] was associated with T1D [82]. A further analysis showed that lncRNAs often regulate genes associated with clusters of islet enhancers and together with transcription factors regulate common genes. One example is lncRNA named PLUTO (PDX1 locus upstream transcript), which regulates a key pancreatic β-cell transcriptional regulator, PDX1 (pancreatic and duodenal homeobox 1). PLUTO controls PDX1 by regulating the 3D architecture of the enhancer cluster in PDX1 locus in human islets. Knockdown of PLUTO is associated with downregulation of PDX1 in primary islet cells and it could be associated with developing both type 1 and type 2 dia-betes [83]. According to a study of immune-mediated disease genes, more than 90% of disease-associated SNPs are placed within non-coding regions of genomes such as ncRNA genes and around 10% of the autoimmune disease associated SNPs are present within lncRNas. The role of lncRNAs in T1D has not yet been profoundly investigated but this is an attractive field to explore. It could help people to better understand the genetic basis for the development of diabetes and to improve diagnostic tests that can detect the disease at its beginning stadium.
MicroRNA
MicroRNAs (miRNAs) are molecules of the size between 18 and 22 nucleotides. They bind to 3’UTR (untranslated regions) of mRNA and cause their degradation or inhibition of translation [84, 85]. Nowadays, more and more studies present miRNA changes in the pathogenesis of diabetes. It has been showed that miRNAs (such as miR-150, miR-146a and miR-424) take part in the formation of autoimmunity and β-cells dysfunctions. Another example are miR-146a NC_000005.10:g.160485411C>G (rs2910164) and miR-155 NC_000021.9:g.25572410T>A (rs767649) variants, which are less common in T1D patients compared to healthy individuals. Moreover, the most common miRNA in pancreatic islets is miR-375 which is a β-cells death biomarker85studies are focusing on other factors that may contribute to the pathogenesis of diabetes, such as epigenetics, a term \”traditionally\” encompassing changes to the DNA that do not alter sequence and are heritable (primary methylation and histone modification. Increase in its amount cause the decrease of β-cells mass. β-cells can release miR-375 into the extracellular environment. This type of miRNA regulates insulin secretion and its expression might be altered in β-cells concomitantly to its circulating level. Furthermore, miR-375 level was downregulated in human pancreatic islets treated with high glucose [86].
Epigenetic modifications in type 1 diabetes
Next to the genetic factors of diseases there are also non-genetic factors which are less clear-cut. These factors are called epigenetic factors and they execute changes in gene expression without altering DNA sequence directly [87]. Epigenetic mechanisms above all include DNA methy-lation and histone modification [85]. Epigenetic modifications control biological processes and are specific to the stage of disease development, which is correlated with the regulation of cell differentiation [84, 85].
DNA methylation
The most common epigenetic alteration is methylation, which is a process based on addition of a methyl group to cytosine at C5 position. It takes place especially when it is followed by guanine, in so called CpG dinucleotides that comprise around 1% of the human genome [84]. DNA methylation usually correlates with gene silencing [84, 87]. By now, 132 positions were identified as T1D-methylation variable positions (T1D-MVPs) and they include HLA class II gene (HLA-DQB1) and GAD2 (glutamate decarboxylase 2) encoding GAD65 autoantigen. T1D-MVPs are changes found in monoclonal zygote twins, therefore they cannot be a result of genetic differences. It is known that some T1D-MVPs are found in individuals before T1D diagnosis. This suggests that they arise very early in the process that leads to T1D and are not simply due to post-onset-associated factors [88]. Moreover, methylation sites also comprise HLA-E, HLA-DOB, HLA-DQ2A, INS, IL-2RB and CD226 as well, which may take part in T1D development. Different levels of methylation of INS genes (Ins1 and Ins2) are correlated with T1D progression and they could be considered another marker for this disease. Moreover, it has been proven that during β-cell destruction associated with T1D onset and progression noticeable changes in methylation (presented by those damaged β-cells’ DNA) are present in the insulin genes accountable for regulation of transcription [85].
Histone modifications
Next group of epigenetic factors are histone modifications. Cell’s genetic material is packed into a chromatin structure (linear DNA is wrapped around a histone core). The N-terminal tails of these histones can be covalently modified [84]. The most common types of these modifications are: 1) histone methylation, 2) acetylation (of lysine and arginine), 3) phosphorylation (of serine and threonine), 4) ubiquitination and 5) sumoylation (of lysine) [84, 85]. The level of lysine 9 (belonging to H3 histone protein – H3K9Ac) acetylation demonstrates an increase in the upstream regions of HLA-DRB1 and HLA-DQB1. And those genetic variants are highly common in T1D patients [85].
Imprinting
Imprinting is another very important epigenetic mechanism. It is based on the transcriptional regulation in which only one parental allele of a specific gene is expressed [85]. Phenotype expression of imprinted genes is a consequence of silencing one allele and allowing a normal state of monoallelic gene expression without altering DNA sequence. It is based not only on sequence variation itself but also on methylation and histone modifications [89]. Some of autosomal loci are actively transcribed only on the maternally inherited chromosome and the others are only transcribed on the paternal chromosome [84]. Imprinting takes part in pathogenesis of many types of diabetes, for example transient neonatal diabetes (TNDM) is caused by abnormalities in the imprinted locus on chromosome 6q24. PLAGL1 gene [PLAG1 (pleomorphic adenoma gene 1) like zinc finger 1] resides in this region and it is normally expressed only on the paternal allele. Duplication of the paternal allele and hypomethylation of the maternal allele cause the overexpression of this gene. PLAGL1 regulates the cell cycle arrest and apoptosis. Its overexpression causes apoptosis and loss of β-cell mass. This finally leads to the loss of insulin secretion [85].
Environmental factors
It is widely known that besides typical epigenetic factors associated with autoimmune disorders, we can additionally distinguish environmental factors which are also capable of affecting disease development. Environmental factors include: 1) temperature, 2) climate, 3) increased hygiene and decreased rates of infection in early childhood, 4) vaccinations, 5) antibiotics and drugs, 6) iodine levels, 7) cigarette smoking as well 8) an increasing wealth. On top of that, diet (especially wheat consumption) also has a great impact on the development of T1D. One of the most important environmental factors are infections. It is proved that bacterial infections play a role in development of pancreatic inflammation. Another type of infections are viral infections, for example myocarditis caused by the enteroviruses family [90]. Possible mechanism of action is based on the viral remnants which can be found in pancreatic islets of patients with T1D and are not present in healthy people. Taking antibiotics and various drugs (especially methyl donors) modifies the composition of intestinal microbiota. It has been shown that reduced microbial diversity, especially low on butyrate-producing and mucin-degrading bacteria, takes part in the autoimmunity of islet cells in children [87].
Conclusions
Type 1 diabetes is a chronic autoimmune disease characterized by islets inflammation and destruction of β-cells, which are responsible for insulin production and regulation of glucose homeostasis. It is very important to understand the molecular mechanisms of T1D development based on genetic studies. GWAS analyses led to the discovery of a number of loci modifying the susceptibility to diabetes. Those loci are considered biomarkers of diabetes development (before and after the onset of the disease) in living organisms. Most of T1D-associated loci encode proteins that are involved in the immune response and inflammatory reactions. The biggest group of genes involved in T1D is present in the human leukocyte antigen region (HLA) that contains almost half of genes associated with T1D. Analysis and identification of novel biomarkers is needed for prevention and monitoring of diabetes development. Future genetic studies are crucial to broaden our knowledge of T1D pathogenesis and find a novel effective therapy. Analysis of a genetic background could result in a discovery of new biomarkers of T1D and in a development of a more specific and sensitive diagnostic test.