
Introduction
Helicobacter pylori is a gram-negative microaerophilic bacterium that is present in the human body both in normal and pathological conditions. It is associated with various diseases of the gastrointestinal tract, such as acute and chronic gastritis, gastric ulcer, gastric lymphoma, and adenocarcinoma [1–5]. Although the bacterium is represented mainly in spiral form, it is capable of forming coccoid forms using the bacillary-coccoid transformation (BCT) process [6].
Since the discovery of these forms, various theories have been proposed regarding the survival, as well as function and role in the pathogenesis of H. pylori-associated gastrointestinal diseases. Despite the fact that significant progress has been made in understanding the morphology and vital functions of H. pylori and the pathogenesis of its diseases, we do not have a clear understanding of the importance of coccoid forms in the development of diseases, nor the importance of identifying them for the selection of rational eradication therapy.
In this article we discuss the most pressing topics that have accumulated over the past 10 years in order to come to an understanding of the importance of coccoid forms, namely:
Coccoid viability
The morphological form of H. pylori is represented by a spiral, under optimal conditions for vital activity. However, the bacterium can turn into a coccoid form under the influence of adverse conditions. This is a round morphological form of H. pylori approximately 0.5–1.4 µm in size, which has even edges and a homogeneous cytoplasm on TEM (transmission electron microscopy) [7]. The presented criteria allow us to distinguish coccoid forms from dead bacteria, which have a similar form. Coccoid forms have a typical bacterial structure, represented by the cytoplasmic membrane, cytoplasm, and undamaged cell wall on TEM [8], which morphologically allows them to be classified as viable forms. Willen et al. revealed a high electron density in the cytoplasm of most coccoid forms, which has one or several vacuoles [7]. It was confirmed that these vacuoles contain a large amount of polyphosphate [9]. Polyphosphates can be used as an energy reserve in case of exogenous energy deficiency, to maintain the integrity of the main bacterial components, such as DNA, RNA, and structural proteins, which indicates the broad adaptive abilities of coccoid forms.
The coccoid forms of H. pylori can be divided into three types according to their structure: the degenerative coccoid form, which is a dying bacterium with a pyknotic structure; alive coccoid bacteria that can be grown in agar medium; and viable but non-cultivable (VBNC) H. pylori [6, 10, 11]. According to other data, the coccoid form is divided into two types: A and B. A type has an irregular shaped, rough surface with few flagella and an indistinct cytoplasmic membrane that obscures much of the intracytoplasmic structure, and these bacteria also tend to adhere to each other and thus are considered to be dead cells of H. pylori. B type shows a smoother surface with tightly encircled flagella, and comparatively clear membrane structure. B type has a better-maintained intracytoplasmic structure and thus is considered as a living cell [12].
Coccoid forms, besides their viability, can also be distinguished on the basis of their metabolic abilities. Sarafnejad et al. [13] showed by using flow cytometry method the presence of metabolic activity of coccoid forms. The study used Rhodamine 123, which absorbs viable microorganisms with greater intensity. Coccoid forms showed the ability to absorb Rhodamine 123 but with less activity than spiral form, which was explained in the study by the fact that this morphological form exhibits less metabolic activity than spiral forms. Furthermore, Loke et al. confirmed that the metabolism of coccoid forms, when compared with spiral forms, is significantly slowed but not stopped. Among proteins that were much less enriched in coccoid forms, 35 were enzymes involved in various metabolic pathways. Conversely, the content of the other 13 metabolic enzymes was increased. Based on a comparison of the relative abundance of key metabolic proteins expressed in two morphologically differentiated forms of H. pylori, coccoid forms were less metabolically active than the spiral forms, but the activity of most enzymes was still maintained at the basal level, while the activity of the others was increased [9].
Slowing down the metabolism is an important mechanism necessary for survival in extreme conditions, such as lack of oxygen, energy substrates, etc. [14]. These data speak in favour of the fact that coccoid form, from the point of view of its metabolic potential, is not only a viable morphological form, but also a special existence strategy that the bacterium chooses to live in adverse conditions. Conversely, the activity of RecA enzyme, which starts the so-called SOS system that participates in the DNA repair of cells, increases [9]. This result is contradictory and is interpreted in two ways. Its increase may be due to damage to the DNA of the bacterium, which leads to death, or to the increased reparative potential of coccoid form, and it facilitates the ability to recombine. One more ambiguous result should be noted, namely a decrease in activity of enzymes (RecN, RecR) that are responsible for DNA repair, the damage of which is caused by oxidative and acid stress [9, 15, 16]. This phenomenon can be caused by an increase in the resistance of the coccoid form to oxygen radicals and acid ions or a decrease of their release due to a slower metabolism and, consequently, less need for DNA repair or having limited reparative ability of the coccoid compared to the spiral forms. An increase in the activity of proteins responsible for chemotaxis, colonisation, and response to “stressful” factors was observed against the background of a decrease in another regulator responsible for the same functions in the same study. The number of structural subunits of urease B (UreB) and auxiliary proteins (UreF) were reduced in coccoid form, which explains the observation of a decrease in the urease activity of this morphological form and failure of urease test in determining the presence of Helicobacter pylori in gastric mucosa. MreB protein, which increases division activity, is actively synthesised in coccoid forms [9]. It is thought that MreB may play a role in the survival of coccoid forms.
The ability to form biofilms, in which the two above morphotypes coexist, also indicates the viability of coccoid forms. Moreover, coccoid forms of H. pylori possess increased capacity to aggregate into monomicrobial bacterial clusters surrounded by a thick matrix of exopolysaccharide [17].
The tolerance of coccoid forms to antibiotics and the human immune system
The increasing H. pylori resistance to previously efficacious antibiotic regimens is of great concern and requires modification of therapeutic strategies [18]. For example, the antibiotic clarithromycin has been shown to be almost completely ineffective in studies in Poland and Turkey [19, 20]. Additionally, H. pylori showed resistance to metronidazole in Polish patients [20]. Thus, it is important to learn and explore the factors that can influence successful eradication treatment of H. pylori infection. Baris et al. and Szadkowski et al. have found different factors that can be involved in antibiotic resistance treatment from genotype, gender, age, smoking habits, and the medication itself, to the resistance of bacteria [19, 20].
One of the reasons for ineffective therapy may relate to antibiotic-resistant coccoid form of H. pylori. Coccoid form of H. pylori usually contributes to antibiotic tolerance and makes the bacterium less accessible for eradication. Figura et al. found that one of the factors negatively affecting eradication was the presence of coccoid forms (p = 0.0008) [21]. Additionally, Kravtsov et al. also showed in their study that the majority of 14 patients whose eradication was ineffective had the highest degree of contamination with coccoid form of H. pylori [22]. In addition, the coccoid form makes recovery more difficult, and causes relapse and epidemic transmission [23]. In a study where 21 biopsies were taken from patients who had eradication, H. pylori was isolated in culture in seven biopsies, while gene expression methods revealed viable H. pylori cells in most biopsies. Scanning electron microscopy (SEM), meanwhile, showed cluster coccoid bacteria located in a microbial biofilm, suggesting that coccoid forms may also contribute to bacterial tolerance to antibiotics used [17]. Faghri et al. [24] tested the sensitivity of viable coccoid forms to three antibiotics and showed that amoxicillin exerted the greatest bactericidal effect in vitro on spiral forms of H. pylori; however, 2 × minimal inhibitory concentration (MIC) did not have a bactericidal effect on coccoid forms. Another study found that coccoid forms were resistant to amoxicillin concentration more than 10-fold compared to the MIC [25], confirming a higher antibiotic tolerance of these morphological forms.
The mechanisms of coccoid resistance to eradication are not well understood, but it can be assumed that bacteria acquire this property by reducing surface area, slowing down metabolism, and expressing “stress genes”. One of these genes is spoT. The spoT gene is a potential global transcriptional regulator for growth and persistence of H. pylori and, apparently, a critical regulator of stress reactions that affects energy metabolism, detoxification, and protein fate [26, 27]. It is believed that the spoT gene is necessary for the survival of H. pylori in the coccoid form under conditions that bacteria may experience during eradication or when in an environment different from living environment [14, 27]. The rapid accumulation of guanosine tetraphosphate (ppGpp) and inhibition of stable RNA synthesis are clear signs of a stringent response. In fact, it is assumed that ppGpp binds and modifies the affinity of RNA polymerase to various promoters, such as those associated with adaptation to adverse conditions, and it has a significant effect on controlling the expression of virulence genes in a wide variety of pathogenic bacteria [28, 29]. In addition, spoT plays an important role in the avoidance of bacteria from macrophage elimination and plays a significant role in colonisation of H. pylori in mice without inducing a strong immune response [26, 27]. Poursina et al. found in his study that expression of the spoT gene was significantly reduced in spiral forms of H. pylori, while the corresponding expression of the gene was increased in coccoid forms [30].
There is also evidence of stability of coccoid forms due to changes in the composition of the cell wall. The fact is that the transition to coccoid forms correlates with the accumulation of the N-acetyl-D-glucosaminyl-β(1,4)-N-acetylmuramyl-L-Ala-D-Glu (GM-dipeptide) motif. This modification may play a role in resistance to bactericidal agents, such as amoxicillin, that affect the cell wall [31]. Costa et al. [32] demonstrated that the activation of (γ)-glutamyl diaminopimelate-endopeptidase during the morphological transition into dipeptide monomers in H. pylori leads to increased conversion that is similar to the sporulation in Bacillus sphaericus. This phenomenon leads to the genesis of resistant coccoid forms and endospores in H. pylori and B. sphaericus, respectively. The high resistance of these forms can be due to various cell wall modifications in coccoid forms and loss of antibiotic targets in these forms.
Moreover, Chaput et al. demonstrated the ability of H. pylori coccoid cells to avoid detection by the immune system due to significant modification of the cell wall peptidoglycan, which did not have IL-8 stimulating activity in the epithelial cells of the stomach. Thus, H. pylori in a state of VBNC may be able to avoid or modulate the host’s reaction and thus persist in the human stomach [31].
We strongly recommend that gastroenterologists accept the H. pylori bacillary coccoid dualism phenomenon and do not persist in prescribing a second course of eradication for coccoid forms of H. pylori. We hope that the BCT of H. pylori will be considered in the next Maastricht agreements.
Bacillary coccoid transformation and its reversibility
As mentioned earlier, the spiral H. pylori can turn into coccoid under the influence of adverse conditions, such as a change in pH, pO2 [14, 33, 34] aging, aerobiosis [35], temperature increase, or antibiotic exposure [24]. Spiral H. pylori can turn into coccoid form, and the conditions must be suboptimal in order to transform into VBNC [11]. However, bacillary coccoid transformation (BCT) can occur in the absence of adverse conditions, for example during prolonged cultivation [36]. There are two possible explanations for this process. According to the first, the concentration of substances that can stimulate BCT increases with prolonged cultivation of bacteria in the colony, thereby increasing the population of coccoid forms [37]. Another explanation is based on the impossibility to simulate the conditions observed in gastric mucosa in vitro, and due to the accumulation of toxic metabolites (when compliant to parameter “suboptimal conditions”), spiral forms turn into coccoid forms over time. This once again confirms that BCT is an active process necessary for the survival of bacteria.
During the process of transformation, H. pylori goes through several stages: S (spiral) form, U form, and C (coccoid) form (Figure 1). These stages are preceded by morphological changes.
Figure 1
Immunocytochemical examination of smears-prints obtained from the antrum of the stomach. A – spiral form, B – U form, and C – coccoid form
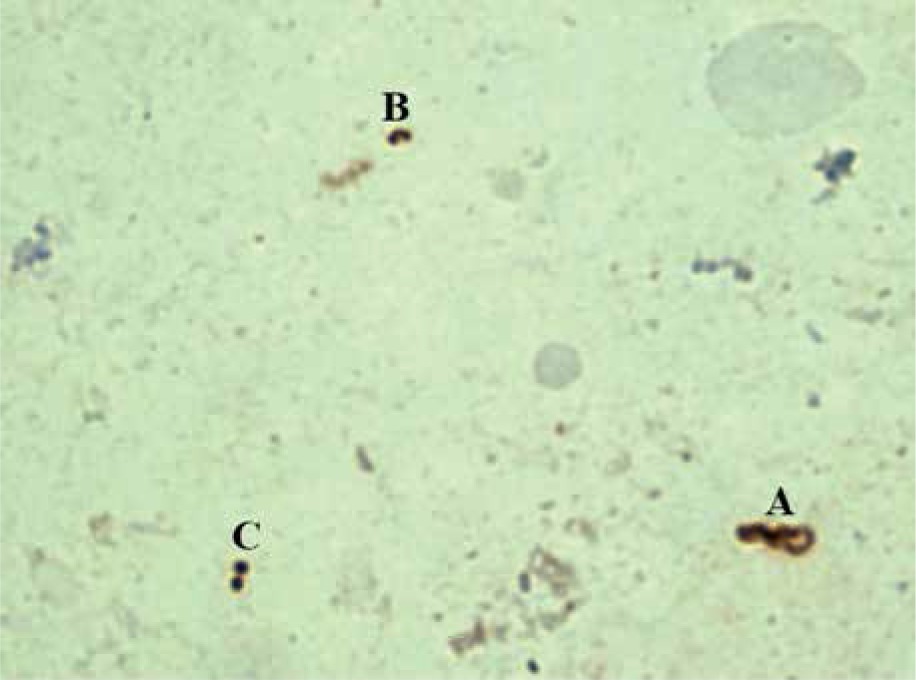
The initial stage of H. pylori transformation into the coccoid form is accompanied by condensation of the protoplasmic matrix and an increase in the periplasm on one side of the microorganism (usually at the pole opposite to the basal complex of the flagellum). The increase in the volume of the periplasmic space leads to stretching of the cell wall, and displacement of the protoplasmic matrix to the periphery and the accumulation of dense material, which leads to the formation of U-shaped cells (U forms) [8]. Furthermore, U forms are transformed into coccoid forms with an increase in the protoplasmic cylinder and formation of a double membrane system [38]. It is worth noting that U forms are only an intermediate stage in the process of turning a helix into a coccoid, and not separate morphological forms [14, 39, 40]. U forms of H. pylori retain the polar membrane associated with the flagellar basal complex [8]. However, not all intermediate forms have a sufficient number of flagella and a sufficient level of metabolic activity, which limits their ability to move [40].
An important fact is that the most active induction of BCT occurs in the most virulent strains [41], which suggest the need to measure the activity of the BCT at least in order to have an idea about the virulence of the bacteria that caused the disease and the possible outcome of the development of the disease.
Clinically significant data suggest that different concentrations of amoxicillin, clarithromycin, metronidazole, and erythromycin, i.e. major AB in the eradication scheme, can induce the formation of coccoid forms [24, 42]. It follows that not all H. pylori microorganisms are completely destroyed after eradication therapy; probably, some of them can be converted into coccoid forms and thus become resistant to antibacterial drugs. This would explain the failure of treatment and the recurrence of infection [43, 44].
The spread of H. pylori in an alimentary tract is also happening due to coccoid forms, which are highly resistant to adverse environmental conditions [45, 46], and possibly contributes to the recurrence of infection [42]. Despite the fact that data were obtained on the transformation of some coccoid forms back into a spiral [47], the ability to reverse the process of coccoid transformation is little studied, but it is not denied that this phenomenon is possible [46]. This process is called “resuscitation” in the literature [48]. In the case of possible return of coccoid to the spiral form, there is a possibility of inducing an active infection on the surface of gastric mucosa.
Therefore, it has been suggested that the transformation of bacteria into coccoid form reduces their metabolic rate and causes minor changes in the physiological processes of the bacterium. These changes, in turn, make it impossible to restore the bacterial signs that contribute to the colonisation and infection of gastric mucosa. The coexistence of both forms (coccoid and spiral) in different environmental conditions in this case explains the survival strategy of the bacterium [49].
The ability of coccoid forms to transmit infection
When the reversibility of BCT in the human body is finally proven, then the question will arise about the possibility of spreading infection using coccoid forms. Krzyżek and Gościniak proposed a hypothetical model for the transmission of coccoid forms of H. pylori and their induction of the infectious process [37]. H. pylori infection occurs by swallowing contaminated water, food, or contact with an infected person. The body is infected mainly by coccoid forms, because the bacterium can exist in this form for long time in the environment [50]. Furthermore, when stimulated by the resuscitation process by the gastric microbiota, which secretes substances like AI-2, coccoid forms turn into mobile spiral forms that colonise the mucous membrane, form biofilms, and contribute to the development of diseases [37]. Subsequently, BCT inductors accumulate in the biofilm, which contribute to the formation of coccoid forms and the release some of them with faeces into the environment [37, 51], ensuring the spread though faecal-oral route. Then, when environmental conditions change (accumulation of BCT inducers, decrease in nutrient levels, etc.), coccoid forms in the stomach turn back into spirals, contributing to the progression of the disease and the search of new niches in the gastric environment.
Virulence and possible role in the development of gastric cancer
The expression of coccoid virulence factors in comparison with spiral forms has not been studied enough, but according to the available data it can be suggested that coccoid forms, if not more, then certainly are no less virulent than spiral forms [9].
Despite the decrease in the BabA factor affecting the adhesive abilities, there are still no significant differences in the index of adhesive abilities between the coccoid and the spiral forms [23].
An increase in the expression of some factors of the cag PAI apparatus shows that the coccoid form is not inert but can infect humans and is likely to play an important role in chronic diseases of the stomach. This fact is confirmed by a clinical case, in which an 89-year-old patient had chronic active gastritis, which was characterised by the presence of only coccoid forms in the coolant [52]. While VacA is an inducer of apoptosis, CagA can lead to excessive cell proliferation [53, 54]. Consequently, an uneven decrease in the expression of the corresponding genes can lead to excessive division or excessive apoptosis. The expression of the cagA and vacA genes was lower in the H. pylori coccoid form than in the spiral form, but the decrease in vacA expression was greater than that of cagA [55]. These results suggest that the abundant contamination with coccoid forms may be one of the factors that increase the risk of developing gastric cancer. The abundance of expression of tumour necrosis factor α (TNF-α)-inducing protein (Hps) with proven carcinogenicity, as shown in Table I [9, 56–63]. Increases in coccoid forms, and also indirectly affects their stability in gastric mucosa, reducing the acidity of the environment [9].
Table I
Expression of H. pylori virulence factors in coccoid forms compared with spiral forms. “+” – greater expression, “–” – lower expression. *The presence of opposite results in different studies i.e. +/- , which may require additional detailed study
| Factor | Expression | Function |
|---|---|---|
| CagA | – [56] | Violation of intracellular signalling systems |
| CagE* | + [9], [57] | Key factors of secretory system type IV. Caga translocation into the cell [59] |
| CagV | + [9] | |
| VacA | – [55] | Induction of vacuolization and apoptosis of cells, reduction of T-lymphocyte activity |
| BabA | – [57] | Adhesion molecule |
| YidC | + [9] | Estimated translocase component |
| Hps | + [9] | Inductor for the expression of TNF and chemokines |
| RocF | – [58] | Hydrolysis of arginine with the formation of urea and ornithine [60] |
| Tip-α | – [58] | Moved to cells, Tip-α induces the expression of TNF-α, IL-6 and several chemokine genes by activating NF-kB [61–63] |
The increased expression of anti-apoptotic protein “survivin” in coccoid forms plays a role in the development of carcinogenesis [23], and possibly increases the risk of stomach cancer formation contaminated with coccoid forms.
Conclusions
Summarising the findings:
based on the data presented above, we assume that coccoid forms are viable morphological forms;
coccoid forms of H. pylori are tolerant to the most effective antibiotics used in eradication therapy;
the question about BCT reversibility requires more extensive research;
coccoid forms of H. pylori may be responsible for the faecal-oral transmission in people, possibly turning back into spirals after successful eradication;
occoid forms of H. pylori are virulent and possibly increase the risk of gastric cancer in patients with abundant BCT activity.
These findings prove the need to evaluate coccoid form of H. pylori contamination of the gut in clinical practice in order to prevent the spread of infection, to prescribe rational antibiotic therapy (change the pattern of eradication therapy), and to clarify the prognosis of the disease.











