






Introduction
Psoriasis is a chronic inflammatory disease affecting approximately 3% of the human population [1]. Its pathogenesis comprises abnormal keratinocyte hyperproliferation and differentiation, dermal vasculature proliferation; and a T cell predominant inflammatory infiltrate of the dermis [2]. The morbid state of the epidermis in psoriasis can be defined regarding excessive mitotic activity and unfinished cellular maturation in which cellular proliferation is too fast for sufficient maturation and manifests as an incomplete synthesis of tonofibrils, and unformed stratum granulosum [3]. The turnover time of the human epidermis, i.e., the transit of keratinocytes from the basal cell layer to the stratum corneum, takes approximately 26–28 days, whereas in psoriatic keratinocytes it is shortened [4]. In normal epidermis, the transit from the basal cell layer to the spinous cell layer takes 13 days, whereas in psoriasis it takes only 48 h [5]. Analogously, the cell cycle of basal keratinocytes lasts 311 h in the healthy skin, whereas in psoriasis it lasts 36 h, which indicates a significant acceleration of keratinocyte proliferation in psoriatic lesions [4]. Cell proliferation requires the synthesis of deoxyribonucleic acid (DNA), essential for cell division [6]. In the production of purines, components of DNA, tetrahydrofolic acid derivatives combine with and transfer one-carbon units into number 2 and 8 positions of the purine base. This process requires the reduction of folic acid to a tetrahydrofolic acid derivative by the enzyme named folic acid reductase [6]. Folic acid antagonists inhibit the production of tetrahydrofolic acid by interfering with folic acid reductase and hindering the synthesis of nucleic acids [6]. Methotrexate, which represents folic acid antagonists, has been known in medicine since the 1950s. The drug, as a more stable and clinically manageable, replaced another folic acid antagonist aminopterin (4-aminopteroyl-glutamic acid). The latter has been known since 1948, when it was found to cause temporary remissions in acute leukemia in children [7].
Methotrexate in the treatment of psoriasis
The usefulness of folic acid antagonists in dermatology was revealed by Gubner et al. who found that aminopterin, which was given to patients with rheumatoid arthritis markedly improved co-existing psoriasis [8]. Methotrexate has been widely used in the treatment of psoriasis since the 1960s, when Ryan et al. reported the effectiveness of methotrexate 2.5 mg administered by mouth daily for 6 days followed by 3 days’ rest [9]. In 1971, Weinstein introduced a new schedule of methotrexate administration for the treatment of psoriasis, based on knowledge concerning the kinetics of keratinocyte proliferation in psoriasis and adjusted chemotherapy with methotrexate with cell-cycle specific action [10]. Since then, methotrexate has been used in small doses (2.5 to 7.5 mg) at 12-hour intervals for a total of three doses at weekly intervals [10]. In the in vitro study of Jeffes et al., human epidermal keratinocyte cell lines, HaCaT (aneuploid immortal keratinocyte cell line derived from adult human skin), and HeLa/D98 (immortal cell line derived from cervical cancer cells), as well as leukocytes – MOLT 4 (T-cell line derived from a patient with T-cell acute lymphoblastic leukemia in relapse) and THP-1 (monocytic cell line derived from a patient with acute monocytic leukemia) were exposed to methotrexate, at concentrations that would be expected to occur in vivo, to mimic the blood levels that result from once-weekly therapy [11]. THP-1 and MOLT-4 were found to be 10–100 times more sensitive to the cytotoxic effects of methotrexate than HeLa and HaCaT cells and above 1000 times more sensitive than primary human keratinocytes. These data indicate that proliferating lymphoid cells are more likely to be a primary cellular target of methotrexate than keratinocytes in patients with psoriasis [11]. However, other tissues, in which there is a high degree of mitotic activity, such as bone marrow, oral and gastrointestinal mucosa, hair matrix, and also psoriatic epidermis are sensitive to interference in nucleic acid synthesis by folic acid antagonists [6]. This interference takes place within hours after the use of methotrexate. Most of the methotrexate is excreted within a few hours, and mitotic inhibition is highest in the hair matrix and intestinal epithelium within 24 h of methotrexate administration [6]. Methotrexate is in about 85% excreted through the kidneys and in 15% via the biliary system in the feces [6]. Regarding the effects of methotrexate on cells, one should also take into consideration the actions of methotrexate on folate-independent pathways. Methotrexate is known to exert effects on mitochondrial oxygen uptake, oxidative phosphorylation and the activity of several enzymes linked to the respiratory chain [12]. Yamamoto et al. revealed that the inhibitory effect of the drug on stage III of the respiration with NAD-linked (nicotinamide adenine dinucleotide-linked) substrates is due to the site of inhibition anterior to the cytochrome b region [12]. It did not affect the ATPase activity of NADH oxidase (E.C. 1.6.99.3), NADH dehydrogenase (E. C. 1.6.99.3), and NADH cytochrome c reductase (E.C. 1.6.2.1) [12]. The study of Singh et al. provided substantial data concerning methotrexate activity. It demonstrated an increase in methotrexate activity in cells with a decreased expression of nicotinamide phosphoribosyltransferase (NAMPT; EC 2.4.2.12) [13]. This action of methotrexate was dependent on its antifolate activity [13].
Nicotinamide-adenine dinucleotide in psoriasis
The enhanced proliferation rate of keratinocytes and lymphoid cells in psoriasis requires an increase in their metabolic activity, cellular respiration, and synthesis of ATP. Cellular respiration comprises three sequential stages: glycolysis in the cytosol, Krebs cycle (citric acid cycle) in the matrix of the mitochondrion, and oxidative phosphorylation in the electron transport chain in the inner membrane of the mitochondrion [14]. Transport of the electrons from the electron donors to the electron acceptors occurs during oxidative phosphorylation. The principal electron donor is a reduced form of nicotinamide adenine dinucleotide (NADH), whereas the final electron acceptor is oxygen [14]. On the one hand, one can speculate that increased proliferation and inflammation in a psoriatic lesion result in NADH rapid consumption. Wollina et al. revealed that indeed the NADH fluorescence, which reflects NADH amount in the skin, was reduced in psoriatic lesions [15]. Unfortunately, this is the only existing study devoted to determining the NADH level in psoriatic skin in vivo [15]. An opposite hypothesis links increased cellular metabolism to the increased NADH/NAD+ ratio. In line with that, Wozniacka et al. demonstrated that after a 4-week application of topical NAD+ in Vaseline ointment administered twice daily resulted in the reduction in erythema, infiltration, and desquamation of psoriatic lesions, and was similar to the effectiveness of 0.1% anthralin [16]. However, there is a lack of comprehensive studies on real-time, in vivo non-invasive monitoring of NADH in psoriatic skin. Recent studies indicate that the biological activity of NADH is not only limited to its role as an electron donor because it was also found to regulate gene transcription [17]. Zhang et al. demonstrated that NADH controlled the transcriptional repressor carboxyl-terminal binding protein (CtBP) [17]. The authors showed that increasing NADH levels stimulated CtBP binding in vivo and enhanced CtBP-mediated repression, which suggests that changes in the nuclear NAD+/NADH ratio allow CtBP to serve as a redox sensor for transcription [17]. Imai et al. revealed NAD-dependent histone deacetylation that connects metabolism and genomic silencing [18]. In this process, NAD+ is a co-substrate in the deacetylation reactions catalysed by sirtuins (silent mating type information regulation 2 homologs) [18]. The deacetylation represents epigenetic silencing and enables blocking of transcription or interference with the maintenance of chromatin boundary regions [19]. Thus, one can assume that alterations of NAD+ and NADH levels in psoriatic keratinocytes can influence transcription and participate in the pathogenesis of the disease. However, such studies have not been conducted in psoriasis so far. Since 1951, when Thoerell and Chance introduced the spectrophotometry method to assess the alcohol dehydrogenase level, more new sophisticated biochemical and biophysical techniques have been developed in mitochondrial NADH evaluation [20]. Nevertheless, the more new methods appear, the more new questions arise. So far, it has been difficult to determine in vivo whether measured NADH fluorescence reflects free NADH or protein bound or both. Moreover, in vivo studies do not differentiate what fraction of NADH is investigated: cytosolic, nuclear or mitochondrial. Concentrations of NAD within the mitochondrial matrix were found to be generally lower than the corresponding measurements performed at the whole HeLa cells [21]. NAD+ and NADH should readily pass through nuclear pores. Therefore cellular perturbations affecting free cytoplasmic levels should also cause changes in the nuclear compartment.
Measurement of nicotinamide-adenine dinucleotide
Technically, measurements of NADH and NADPH (nicotinamide adenine dinucleotide phosphate) are indistinguishable, so the sum of both molecules is assessed [20]. The response of NADH to the metabolic transition from normoxia to hypoxia is more complex than a simple increase in NADH concentration [22]. Spectrophotometry methods utilize excitation of the NADH molecule with ultraviolet radiation (UV) of 330 nm, and subsequent emission of fluorescence with a peak at the range of 440–450 nm, which is blue light. In contrast, NAD+ is not fluorescent [20]. Since the introduction of new methods, it has become possible to record changes in NADH continously levels in mitochondria of various tissues and organs in situ [23]. Regarding the real-time measurement of NADH fluorescence in the epidermis in vivo, the AngioExpert device is well suited for such an assessment. AngioExpert makes it possible to assess not only the baseline level of NADH fluorescence, but it can also measure changes in NADH fluorescence of the forearm skin in response to ischemia caused by inflation of a brachial cuff, and subsequent hyperemia following deflation of the brachial cuff [24]. Since maximum light penetration of 340 nm is only about 0.3 to 0.5 mm in the skin, measured NADH fluorescence reflects the NADH amount in the epidermis [24]. A typical epidermal blue fluorescence of NADH in the forearm of a 27-year-old woman during rest in a seated position, then 100-second ischemia and subsequent hyperemia is shown in Figure 1.
Figure 1
A typical epidermal blue fluorescence of NADH in the forearm of a 27-year-old woman during rest in a seated position, then 100-second ischemia and subsequent hyperaemia
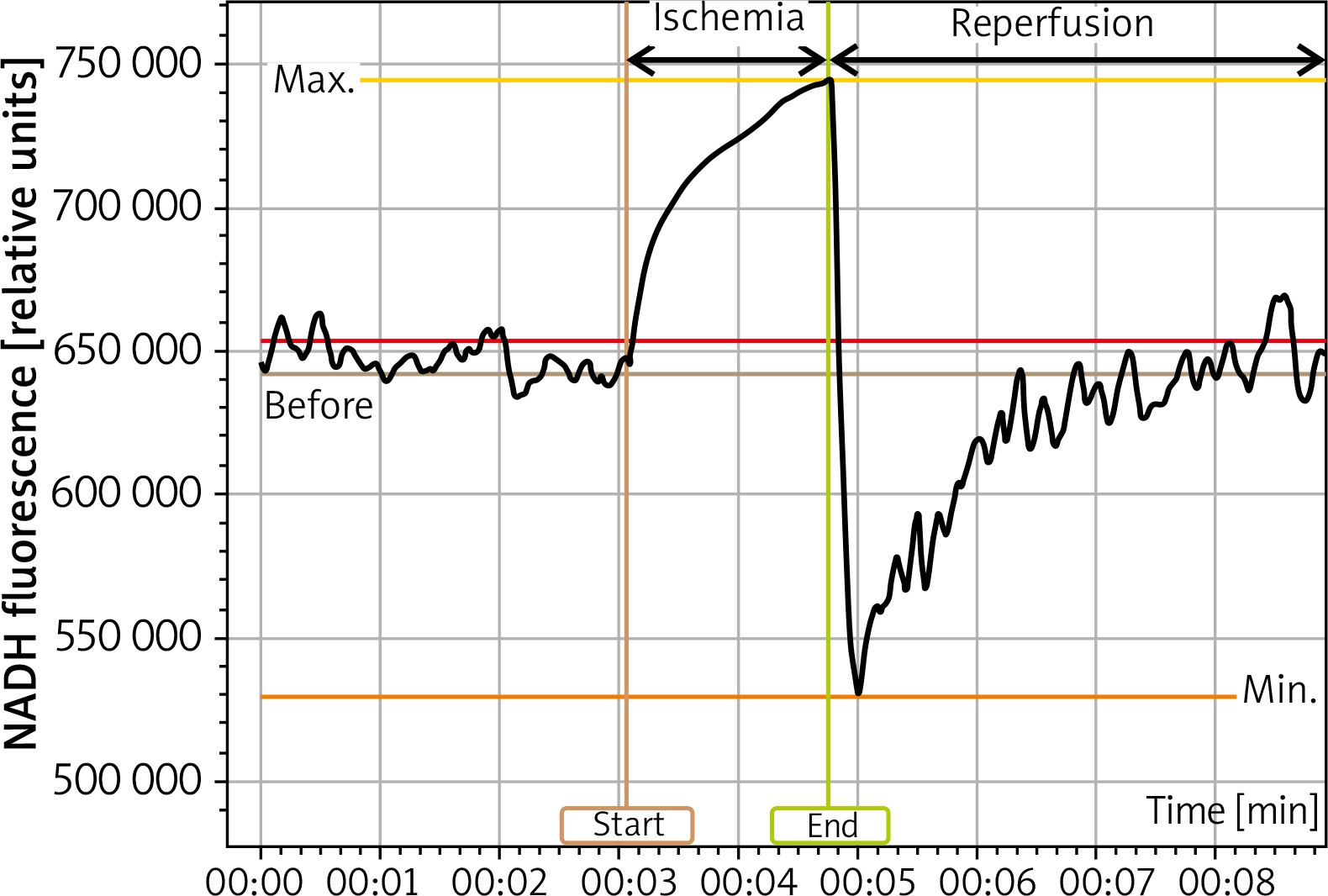
The fluorescence rises in response to 100 seconds of ischemia as a result of the accumulation of NADH in keratinocytes, which is displayed as IRauc (Ischaemic Response area under the curve). Fluorescence rapidly decreases within the first few seconds of reperfusion after deflation of the brachial cuff and recovers within several minutes less steeply, which is displayed as a HRauc (hyperemic response area under the curve).
Conclusions
The data mentioned above emphasize the fact that with the advent of new technologies used for monitoring of NADH fluorescence, more aspects of physiological and pathological conditions of the human skin are elucidated, provide a more comprehensive understanding of the pathogenesis of psoriasis. However, there are still some limitations on such approaches, which require further studies in this field. Undoubtedly, there are insufficient data regarding the role of NAD+ and NADH metabolism in keratinocytes in patients with psoriasis. Moreover, the evidence supporting the hypothesis that methotrexate influences such metabolism is lacking.