
Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is responsible for causing COVID-19 disease (coronavirus disease 2019), which since December 2019 has affected over 173 million people around the world, and caused death of nearly 4 million. The COVID-19 pandemic is responsible for personal, psycho-emotional, moral and economic losses [1, 2].
COVID-19 infection most often occurs by droplet contact with an aerosol containing virions. This virus shows the greatest affinity to the cells of the respiratory epithelium of the lower respiratory tract and the gastrointestinal tract, which results from the necessity to bind to the membrane angiotensin-converting enzyme 2 (ACE2) receptor. Interestingly, the extension of tissue tropism, e.g., to the pharyngeal mucosa, indicates the presence of a furin-like protease, which is necessary for virus entry into the host cell in the upper respiratory tract. The membrane receptor for angiotensin converting enzyme 2 is the binding site of the RBD domain within the S1 protein of the virus, and through a cycle of conformational changes involving the S2 protein this connection allows the virus to penetrate the cell [3–6].
The viral genetic material is released in the cytosol – a single strand of ssRNA demonstrating a positive polarity (+). After the translation of basic proteins, the host’s ribosomes form a reverse transcription complex (RCT), where the viral genetic material is multiplied. Within the viral RNA, non-structural (necessary for replication), structural and auxiliary proteins are encoded. From the clinical point of view, the most important are 4 basic structural proteins, i.e., protein S (spike) – a fusion protein responsible for binding to the host cell receptor, protein E (envelope) – a coat protein responsible for the formation of virions, protein M (membrane) – a membrane protein, the main protein of the viral matrix, and the N (nucleocapsid) protein – a nucleocapsid protein that protects the RNA of the virus [7, 8].
Antibodies directed against the S1 protein and the N protein play a key role in immunological diagnostics. After contracting COVID-19, specific immunoglobulins for these both proteins are produced, while after vaccination, antibodies against the N protein do not occur, due to the fact that the vaccine mRNA does not contain the N protein gene. In the genetic vaccines available on the market (Pfizer, Moderna) there is mRNA encoding only the S protein. And specific antibodies against this protein only are produced in vaccinated persons [9, 10].
The term “cytokine storm” is directly associated with the notion of severe COVID-19 by most clinicians. The influence of individual interleukins and their individual configurations in relation to each other may show both pro-inflammatory and silencing effect on the inflammatory process. Among the whole range of different cytokines, 3 were selected, which were subjected to more detailed analysis, i.e., interleukin-8 (IL-8), interferon-γ(IFN-γ) and transforming growth factor-β (TGF-β).
Interleukin-8 (IL-8, CXCL8) is the most potent chemokine in the human immune system, acting through specific receptors on cells of the immune system to stimulate their migration to the target site. IL-8 has the most potent influence on neutrocytes, monocytes and B cells. Additionally, this factor enhances neutrophil adhesion to the vascular endothelium and mediates angiogenesis [11–13].
IFN-γ is a glycoprotein representing the entire group of interferons, i.e., factors produced in response to the presence of intracellular pathogens, the name of which comes from its ability to interfere with the replication process of viruses inside the host cell. The pleiotropic effects of IFN-γ include among others enhancement of antigen presentation, arrangement of the activation of the innate immune system, coordination of interactions between lymphocytes and endothelium, regulation of Th1/Th2 balance, as well as control of cell proliferation and apoptosis [14, 15].
TGF-β also has a pleiotropic effect. The type of effect caused depends on the type of cell it is acting on and its stage of development. Most generally, it is responsible for the control of cell growth, proliferation, differentiation and apoptosis, and thus the physiological programmed death [16, 17].
Aim
The aim of the present study was to assess the impact of the epidemiological status, selected demographic parameters and the concentration of IL-8, IFN-γ and TGF-β on the post-SARS CoV-2 vaccination response in patients vaccinated with the mRNA vaccine (Pfizer/BioNTech).
Material and methods
In the study conducted among healthcare professionals from November to April 2021, 47 adult volunteers participated, who were vaccinated against SARS-CoV-2 with Comirnaty mRNA vaccine (Pfizer/BioNTech).
In all patients, 3 study visits to the Clinic were planned. The first one before qualification for vaccination, and other two 3 weeks after the first and second dose of the SARS CoV 2 mRNA vaccine, respectively.
During each visit, a detailed medical history was collected, subjects filled questionnaires regarding their current health status, exposure to SARS-CoV-2 infection, and at the second and third visits, they additionally assessed symptoms occurring after two doses of the vaccine. In addition, during these 3 visits, blood was collected from the vein in the ulnar fossa in order to determine levels of selected parameters of the humoral response: anti-SARS-CoV-2 antibodies (for the nucleocapsid phospholipoprotein and the S1 subunit of the spike protein) and cytokines IL-8, TGF-β, IFN-γ.
All patients gave their written consent to participate in the study. The study protocol was approved by the Ethical Committee of Collegium Medicum in Bydgoszcz (KB 161/2021).
Specimen collection
The material for the study was venous blood serum. Blood was taken in accordance with the standard conditions, between 7.00 am and 9.00 am. The patients remained on fasting after an overnight rest. Blood was collected from the median cubital vein using a closed vacuum system (Vacuette, Greiner Bio-One), into a “CAT Serum Sep Clot Activator” 5 ml tubes. The blood samples, after complete clotting, were centrifuged for 15 min at 3500 rotations per min. Serum was immediately separated and kept deep frozen, at –70°C, until assayed.
Laboratory analysis
Anti-SARS-CoV-2 IgG
The concentration of antibodies against two antigens of the SARS-CoV-2 virus in the IgG class was measured by the “Polycheck Infection Serology Anti-SARS-CoV-2 Immunoassay” test. The Polycheck Anti-SARS-CoV-2 IgG assay is an enzyme immunoassay for the quantitative measurement of antigen-specific immunoglobulin G (IgG) in serum to Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). This test is useful as a diagnostic tool to identify individuals with adaptive immune response to SARS-CoV-2. The Polycheck assay used two recombinant proteins representing nucleocapsid phosphoprotein (N) and the S1 subunit of the spike protein (S) of the virus. The use of these two antigens in one single test allows the simultaneous measurement of the concentration of antibodies specific for two SARS-CoV-2 antigens. The antigens as well as calibrators are coated separately in lines on a carrier which is fixed in the well of Polycheck cassette. The results are displayed in arbitrary units (kU/l). Values can either be interpreted as non-reactive (< 0.35 kU/l), grey zone (0.35–0.7 kU/l; repeat measurement recommended) or reactive (> 0.7 kU/l; reactive levels at least one of two antigens can be considered as immunoreactivity against SARS-CoV-2). The Polycheck Infection Serology Anti-SARS-CoV-2 Immunoassay showed a sensitivity of 98.6% and a specificity of 98.7%. Cross-reactivity with common coronaviruses has been excluded by the manufacturer of the test.
Cytokines
The concentration of serum IL-8, IFN-γ and TGF-β1 was measured by commercial available ELISA kits (PharmaGenie ELISA Kits).
During the assay procedure the capture antibody highly specific for target cytokines has been coated to the wells of the microtiter strip plate provided during manufacture. Binding of the appropriate cytokine from samples and known standards to the capture antibodies and subsequent binding of the biotinylated anti-target cytokine secondary antibody to the analyte is completed during the same incubation period. Any excess unbound non-specific analyte and secondary antibody is removed. The HRP conjugate solution is then added to every well including the zero wells, following incubation excess conjugate is removed by careful washing. A chromogen substrate is added to the wells resulting in the progressive development of a blue coloured complex with the conjugate. The colour development is then stopped by the addition of acid turning the resultant final product yellow. The intensity of the produced coloured complex is directly proportional to the concentration of target cytokines present in the samples and standards. The absorbance of the colour complex is then measured and the generated OD values for each standard are plotted against expected concentration forming a standard curve. This standard curve can then be used to accurately determine the concentration of target cytokines in any sample tested.
The Human (IL-8, IFN-γ or TGF-β1) PharmaGenie ELISA Kits (Human IL-8 ELISA Genie, Human IFN-β ELISA Genie, Human TGF-β1 ELISA Genie) are ELISA kits for the detection of IL-8, IFN-γ or TGF-β1 in a range of sample types. These tests are a solid phase sandwich ELISA for the in-vitro qualitative and quantitative determination of IL-8, IFN-γ or TGF-β1 in supernatants, buffered solutions or serum and plasma samples. These assays recognize both natural and recombinant human IL-8, IFN-γ or TGF-β1 without any cross reactivity with other human cytokines in individual tests. The declared sensitivity of these kits are 18.18 pg/ml for IL-8, 9.375 pg/ml for IFN-γ and 18.75 pg/ml for TGF-β1. Maximum measurable concentration IL-8 is 2000 pg/ml, IFN-γ is 400 pg/ml and TGF-β1 is 2000 pg/ml.
All measurements were performed in one series according to the protocols included in the test kits. The INFINITE F50 (TECAN) ELISA microplate reader with the dedicated Magellan for INFINITE F50 software was used to read the absorbance and calculate the results.
Statistical analysis
The comparison of values of quantitative variables in measurements repeated three and more times was performed using the Friedman test. After detecting statistically significant differences, post-hoc analysis (Wilcoxon’s paired tests with Bonferroni correction) was performed to identify statistically significantly different measurements. The comparison of values of quantitative variables in two groups was performed using the Mann-Whitney test. Correlations between quantitative variables were analysed using the Spearman correlation coefficient. A significance level of 0.05 was adopted in the analysis. Thus, all p-values below 0.05 were interpreted as showing significant relationships. The analysis was performed in R software, version 4.1.0 and Microsoft Excel 365.
Results
Forty-seven patients (39 women and 8 men) aged 25–72 (mean: 47.3 ±11.2) participated in the study. The general characteristics of the study population are presented in Table 1.
Table 1
General characteristics of the study population
In the study population, 21 (43.48%) people had normal body weight, defined as body mass index (BMI) < 25 kg/m2, 17 (36.96%) people were overweight, and 9 (19.57%) people were obese.
In all patients the concentration of IgG specific for nucleocapsid phosphoprotein (N) and the S1 subunit of the spike protein (S) was determined at 3 time points.
The first determination was done prior to the vaccination, and other two were done 3 weeks after the first and the second dose of the vaccine, respectively. The study population included 18 COVID-19 convalescents and 29 SARS-CoV-2 naïve subjects.
The concentration of SARS-CoV-2 antibodies in the study population is presented in Table 2.
Table 2
Phosphonouclein-specific IgG concentrations of nucleocapsid and subdivision spike S1 before vaccination and after the first, and second dose
| Variable | IgG nucleocapsid phosphoprotein | P-value | IgG spike S1 | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Before vaccination | After first dose | After second dose | Before vaccination | After first dose | After second dose | ||||
| Patients (n = 47) | Min. | 0.18 | 0.19 | 0.17 | 0.17 | 0.22 | 32.00 | ||
| Max. | 75.00 | 53.00 | 69.00 | 91.00 | 260.00 | 435.00 | |||
| Mean | 10.88 | 7.34 | 9.54 | 0.065 | 6.19 | 65.30 | 196.11 | < 0.001* | |
| ± SD | 21.17 | 12.45 | 16.62 | 15.82 | 56.38 | 94.85 | |||
| Female (n = 39) | Min. | 0.18 | 0.19 | 0.17 | 0.17 | 0.22 | 32.00 | ||
| Max. | 75.00 | 47.00 | 69.00 | 91.00 | 184.00 | 435.00 | |||
| Mean | 0.27 | 0.23 | 0.24 | 0.19 | 1.50 | 95.00 | |||
| ± SD | 22.73 | 11.23 | 17.30 | 17.14 | 49.59 | 98.77 | |||
| Male (n = 8) | Min. | 0.27 | 0.23 | 0.24 | 0.19 | 1.50 | 95.00 | ||
| Max. | 23.00 | 53.00 | 38.00 | 11.00 | 260.00 | 320.00 | |||
| Mean | 3.77 | 7.71 | 9.04 | 1.70 | 75.79 | 197.63 | |||
| ± SD | 7.32 | 17.18 | 12.77 | 3.53 | 80.96 | 72.75 | |||
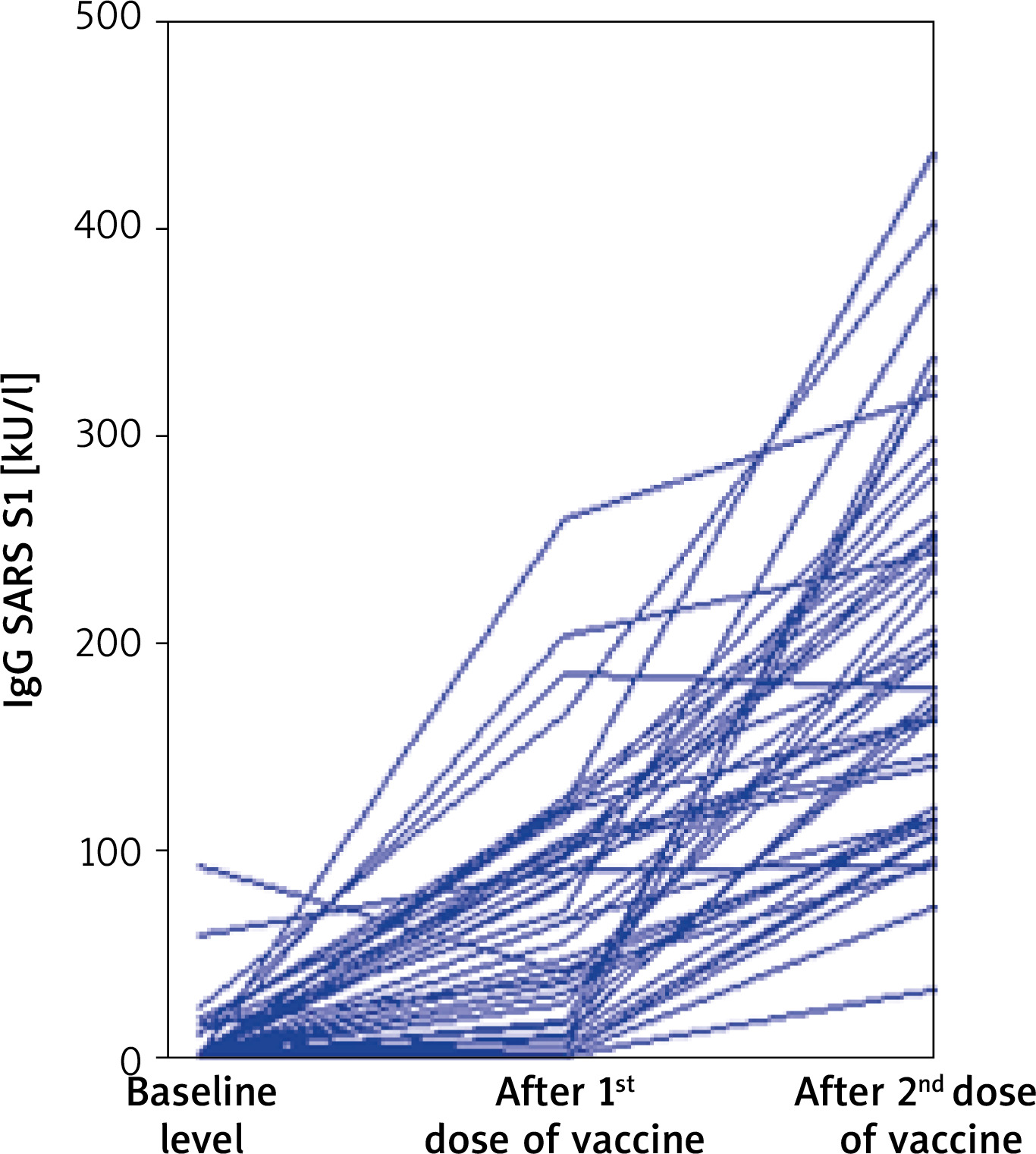
A statistically significant increase in the concentration of IgG SARS S1 was observed in subsequent measurements (Figure 1).
Figure 1
Parallel coordinates plot for each patient’s IgG SARS S1 before vaccination and after first and second dose
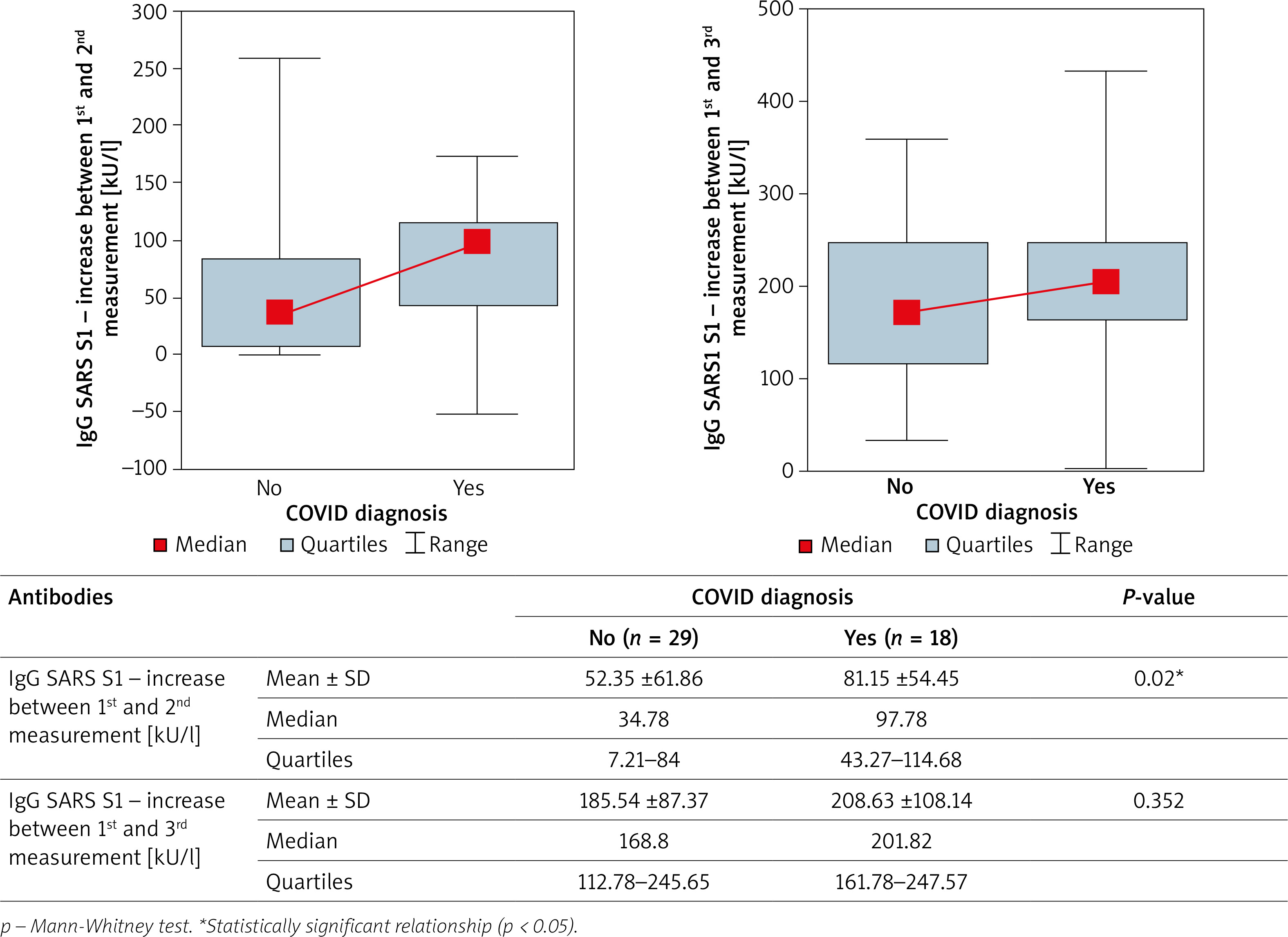
The concentration of IgG antibodies specific for both SARS-CoV-2 proteins in convalescents and people who did not suffer from COVID-19 was analysed. The results are shown in Table 3. Statistically significantly higher levels of S1 spike protein-specific IgG were found after the first dose of the vaccine in COVID convalescents. The effect of COVID-19 on the concentration of sIgG S1 antibodies showed no statistical significance for the IgG S1 concentration after the second dose of the vaccine (Figure 2).
Table 3
Concentrations of IgG specific to nucleocapsid phosphoprotein and subdivision spike S1 before vaccination and after first and second dose of mRNA vaccine
Figure 2
Influence of positive COVID diagnosis before vaccination on the rise of antibodies after first and second vaccination
There was no statistically significant effect of age, BMI and gender on the increase in antibody levels after the next two doses.
No statistically significant increase in the concentration of the determined cytokines (IL-8; TGF-β and IFN-γ) was demonstrated in subsequent determinations before vaccination and after the first and second dose. Most of the studied patients showed low concentration of serum IL-8 and IFN-γ, below the sensitivity of the immunoassay method used (Table 4).
Table 4
Concentration of IL-8; IFN-γ and TGF-β before first vaccination, after first and after second vaccination. In most cases, IL-8 and IFN-γ was below the sensitivity of the test
| Cytokine | Before vaccination | After first dose | After second dose | P-value | |
|---|---|---|---|---|---|
| IL-8 [pg/ml] | Mean ± SD | 21.14 ±12.32 | 20.81 ±11.73 | 20 ±7.07 | 0.024* |
| Median | 18.78 | 18.78 | 18.78 | ||
| Quartiles | 18.78–18.78 | 18.78–18.78 | 18.78–18.78 | ||
| IFN-γ [pg/ml] | Mean ± SD | 23.11 ±92.27 | 24.73 ±103.6 | 18.02 ±57.51 | 0.948 |
| Median | 9.38 | 9.38 | 9.38 | ||
| Quartiles | 9.38–9.41 | 9.38–9.38 | 9.38–9.38 | ||
| TGF-β [ng/ml] | Mean ± SD | 3.41 ±0.84 | 3.59 ±1.3 | 3.38 ±1.05 | 0.108 |
| Median | 3.16 | 3.31 | 3.19 | ||
| Quartiles | 3–3.66 | 2.99–3.74 | 2.92–3.61 | ||
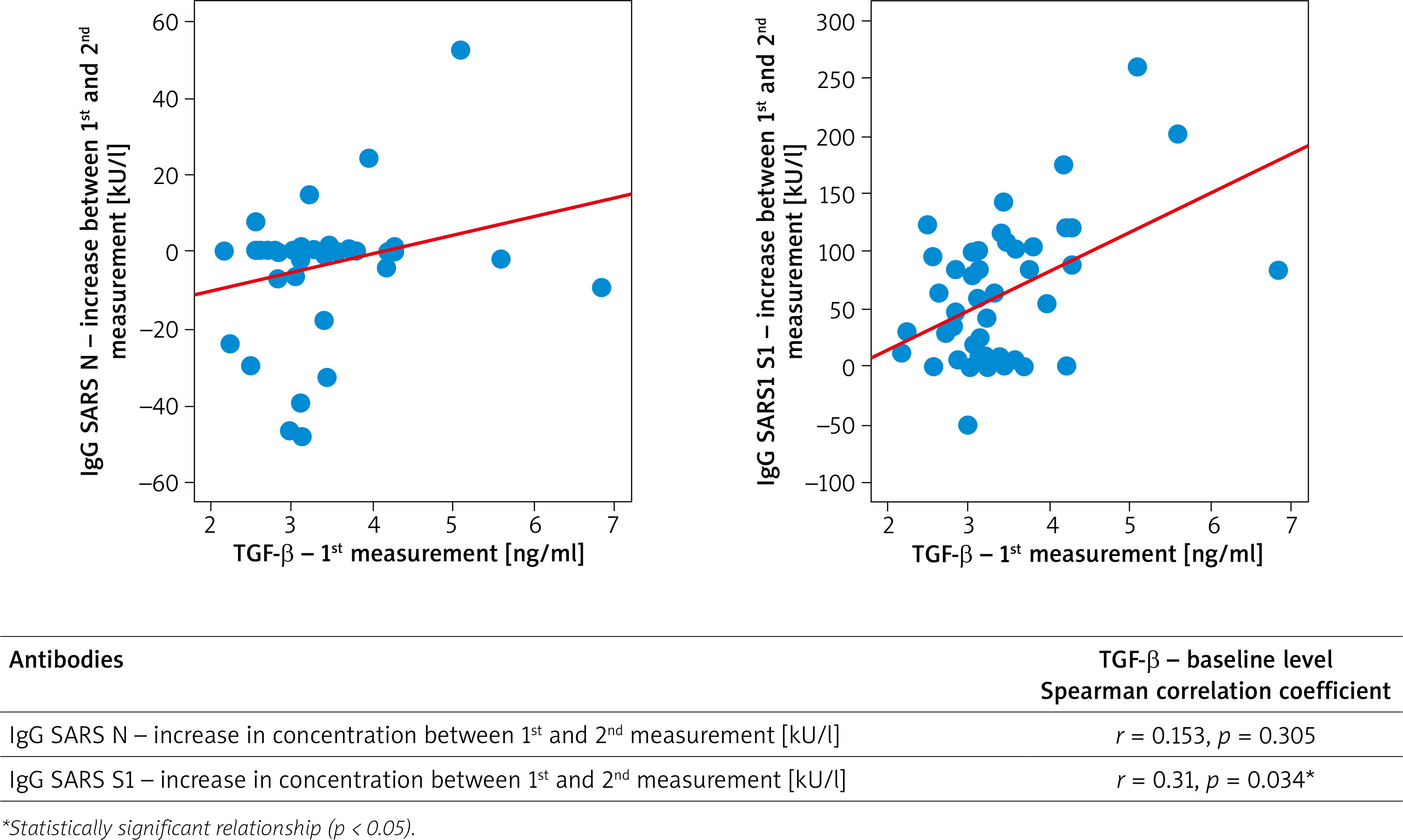
The epidemiological status (convalescence versus a person who had had no COVID-19) had no effect on the baseline serum TGF-β concentration (Table 5). Interestingly, there was a correlation between the concentration of TGF-β and the increase in the concentration of antibodies after vaccination. However, it was found that the initial TGF-β concentration correlated significantly (p < 0.05) and positively (r > 0) with the increase in SARS S1 IgG after administration, the higher the initial TGF-β concentration, the greater the increase in SARS S1 IgG after administration of the first dose of the vaccine (Figure 3). This effect was not observed after the second dose of the vaccine.
Table 5
Influence of COVID in past medical history on the baseline concentration of IL-8, IFN-γ and TGF-β
Discussion
The COVID pandemic has left a significant mark on people’s lives around the world. It caused social isolation, collapse of many enterprises, and the global economic crisis. Most of all, however, it cost lives of many people. Working on a vaccine to provide immunity or alleviate the severity of COVID-19 has become a primary goal of pharmaceutical companies from the initial stages of the pandemic. The use of modern and innovative techniques in the field of genetics, biomedical engineering and microbiology gave hope for quick access to the expected “remedy” for the epidemic. The essence of “genetic” vaccines approved for use is the introduction of fragments of the genetic material of the SARS-CoV-2 virus into host cells. This genetic material encodes molecules showing the highest immunogenicity. Presentation of the antigen, after its direct translation from the introduced mRNA, by antigen presenting cells (APCs), stimulates cells of the immune system to response and to generate specific neutralizing antibodies against the original molecule. By assumption, these specific antibodies, by blocking a selected fragment of the SARS-CoV-2 virus, prevent its penetration into the host cells by inhibiting connection with the ACE 2 receptor [18]. As always, theoretical assumptions must still be supported by hard scientific evidence for the product to be approved for human use. Further difficulties standing in the way of producing an effective vaccine were: lability of the mRNA material, prevention of pre-translational intracellular processing of the introduced molecule, and selection of a “package” that would protect against damage and at the same time allow penetration into the cell through endocytosis [19, 20].
The Pfizer vaccine BNT162b2 (Comirnaty) was first approved by the Official Medicines Agencies, and then the Moderna preparation was also approved. Both vaccines share a similar mechanism of action and demonstrate similar efficacy.
All subjects were vaccinated with the Pfizer preparation, which resulted directly from official arrangements of the Polish government and the Ministry of Health, and at that time it was also the only available option as other preparations were brought to the Polish pharmaceutical market much later [21]. All participants in the study received two doses of the vaccine in accordance with the Summary of Product Characteristics [22].
The subjects developed a specific post-vaccination response in the form of production of sIgG antibodies against the S1 protein of the SARS-CoV-2 virus. In addition, a statistically significant increase in the concentration of specific immunoglobulins against the S1 protein was observed in all of them after the first and second dose of the vaccine. These results are consistent with the data presented in analyses evaluating the efficacy of vaccination against COVID-19 [18, 23–25]. However, it should be noted that the concentration of specific antibodies is not always associated with resistance to infection. Complexity of the immune system does not always suggest that the higher the level of immunoglobulins, the greater the resistance to infection. Attention should also be paid to the individual immunoglobulin subclasses, their specificity, the degree of affinity to the antigen, and finally to other, equally complex part of immunity, i.e., the innate cellular response. Therefore, when assessing the effectiveness of a given vaccine, most researchers use a decline in morbidity, likelihood of severe disease symptoms or death as endpoints rather than serum concentration of specific antibodies [18, 26–28]. Nevertheless, a gradual increase in the concentration of IgG S1 may suggest an increased activity of the humoral response and enhancement of the immune memory associated with both antibody production and cellular response after the next dose of the vaccine [29]. This enhancement occurs regardless of the possible primary COVID-19 incidence, age, gender or BMI.
In the conducted study, IgG antibodies to the N protein of the SARS-CoV-2 virus were also determined. The concentrations of these immunoglobulins showed no relation to vaccination, which is understandable in view of the mechanism of action of the vaccine. The genetic material encoded in the vaccine mRNA does not contain the gene encoding the N protein, therefore this protein is not produced during translation [30]. Due to the above, the presence of antibodies to the N protein indicates the disease of COVID-19. These immunoglobulins gradually decline over time after exposure. Both decreases and increases in IgG concentration for the nucleocapsid protein were observed in the subjects. Moreover, the increased concentration of IgG N was also observed in some people who were considered naïve to COVID, due to the fact that the presence of viral RNA in the mucous membranes of the upper respiratory system was never detected by RT-PCR. The most probable cause of this phenomenon seems to be re-exposure to the virus, which one more time activates the production of antibodies, similar to other viral diseases, or asymptomatic/mildly symptomatic infection, which has never been objectively confirmed by RT-PCR results [31]. At the time when the material for laboratory determinations was taken, most participants were exposed to periodic exposure to SARS-CoV-2, due to the high incidence at that time in the study population, as well as due to the medical profession performed, which applies to all subjects [32].
Many publications indicated a correlation between the severity of COVID-19 and the coexistence of other comorbidities. A correlation between the BMI value and poor prognosis in the course of COVID-19 seems to be interesting. It was pointed out that the increase in BMI value associated with the overweight and obesity of a given patient showed a positive correlation with a more severe course of COVID-19 and significantly reduced the prognosis for a positive endpoint [33–35]. In this analysis, a relationship between many independent factors and the level of IgG S1 antibodies was sought. In the obtained results, BMI did not correlate with the antibody concentration after the first and the second dose of the vaccine. Pellini et al. observed a negative correlation between BMI and the level of specific antibodies after vaccination with the same preparation [36]. This discrepancy may result from the size of the studied population (248 vs. 47), the method used to determine antibody concentrations or differences resulting from environmental factors, i.e., climate, lifestyle, eating habits, physical activity, etc. The authors are inclined to assume that overweight and obesity (defined as BMI > 25 kg/m2 or BMI > 30 kg/m2, respectively) are theoretical grounds for impaired post-vaccination response, and therefore for decreased antibody levels after vaccination in obese subjects. Similar data indicating an impairing effect of obesity on response to the vaccine were obtained during the analysis of the efficacy of vaccines against rabies, tetanus or influenza A/H1N1 [37–39]. Some postulated factors linking obesity with the post-vaccination response include: hormonal activity of adipose tissue, which through adipokines has the potential to bind metabolic processes with the immune system, and an increased risk of obesity-related lifestyle diseases that indirectly affect the immune system.
Age and sex of vaccinated subjects were also among other analysed factors influencing the concentration of antibodies after vaccination. As in the case of BMI, it was not possible to establish a statistical relationship between the above-mentioned factors and the concentration of specific IgG S1. Pellini et al. found that women, thin and young people were characterized by the ability to elicit a more effective humoral immune response compared to men, overweight people and the elderly population [36]. In the current analysis, these relationships have not been confirmed, but certainly studies on a larger population are crucial to finally determine whether mRNA vaccines are highly effective regardless of age, sex, body weight, and comorbidities.
This study confirmed that the first dose of the mRNA vaccine caused a greater increase in antibody concentration in people who have had SARS-CoV-2 infection, compared to those who did not. The pandemic causes huge losses in terms of loss of human life and long-term complications of COVID-19 disease. Moreover, economic effects of the introduced sanitary restrictions are dramatic. There is no doubt that achieving the herd resilience of a population as quickly as possible is a priority. Due to the initial lack of a sufficient number of vaccines in relation to the needs, the authorities of various countries introduced priority vaccination groups, e.g., medical personnel, the elderly, people with diseases predisposing them to a severe course of COVID-19 [40]. One of the ideas for increasing the availability of vaccinations to a wider population was to vaccinate convalescents with 1 dose of BNT162b2 (Pfizer), which was originally a two-dose preparation. In Poland, this project, although discussed publicly by politicians and supported by a positive opinion of the Supreme Medical Council, never entered into force [41]. Such an approach certainly has some disadvantages, it was primarily burdened with the awareness that the vaccination would be administered outside the scope of the drug registration. On the other hand, it made it possible to redistribute the second dose to people at risk awaiting vaccination [42]. Results of the study by Gobbi et al. indicate that administration of a single dose of mRNA vaccine was highly effective in convalescents, with antibody concentrations after the first dose comparable to that achieved after the second dose by those who had been COVID-naïve [43]. In our study, the concentration was not the same, as the average IgG S1 concentration after the first dose in convalescents was 95.08 kU/l, and in COVID-naïve subjects it was 179.69 kU/l after the second dose. At the same time, the concentration of antibodies after the second dose in convalescents remained slightly higher (Table 3). An important conclusion from the present study is that the administration of the second dose equates, in a way, the convalescents to the COVID-naïve subjects in terms of vaccine response. Another aspect of the post-vaccination response to consider is that immunity in convalescents is not necessarily related to IgG S1 levels. A person who has contracted COVID-19 has been exposed to a full range of antigens, which can cause the production of other types of antibodies as well. Moreover, in convalescents there is not only humoral immunity, but also cellular immunity, dependent on T lymphocytes. Moreover, cellular immunity after vaccination is also important, and the analysis of antibody concentration alone is insufficient to assess who is at higher risk of reinfection [44].
When analysing the problem of a pandemic, two positions should be taken into account, which unfortunately do not always coincide. They are a benefit to the individual and a benefit to the population. Based on the analysis of the population effect of delaying the second dose of vaccine, which was performed by Romero-Brufau et al., it can be concluded that such a procedure, especially in the population < 65 years of age, brings population benefit with insufficient vaccine supply [45]. At the same time, from the point of view of an individual, completing the full vaccination course allows for obtaining higher concentrations of neutralizing antibodies, is in line with the drug registration recommendations and offers a potentially better protection against the disease. The pandemic of the present scale generates many ethical dilemmas that are difficult to solve.
To our current knowledge, the present study is the first to analyse the effect of serum concentrations of TGF-β at the baseline on the response to SARS-CoV-2 vaccination with the S1 protein specific mRNA vaccines. One of the most key observations from the present study is the fact that subsequent doses of the vaccine did not statistically significantly affect IL-8 levels; TGF-β and IFN-γ as subsequently measured before vaccination and after the first and second doses. Levels of the cytokines IL-8 and IFN-γ were very low in all determinations. However, baseline TGF-β levels were found to correlate significantly and positively with the increase in SARS S1 IgG after the 1st dose of the vaccine, and this effect was not related to the epidemiological status of the patient. TGF-β is a known tissue repair stimulant. It also has an immunosuppressive effect. It is a strong tissue growth regulator, which may be used in some diseases, for example, increasing its activity accelerates wound healing [46]. This beneficial effect of TGF-β is also crucial in the development of severe diseases such as benign prostatic hyperplasia, scleroderma, pulmonary fibrosis, glomerulosclerosis, tubulointerstitial fibrosis, liver fibrosis, and open angle glaucoma. In the context of COVID-19, there are currently several theoretical models that postulate that inhibition of TGF-β activation pathways may have a beneficial effect on prevention of late complications of SARS-CoV-2 infection, such as pulmonary fibrosis in particular. It is also known that SARS-CoV-2 infection causes an increase in the serum concentration of TGF-β [47]. TGF-β also inhibits proliferation of B cells, and also promotes apoptosis of their immature or naive forms, while promoting proliferation and differentiation of Langerhans cells. It may also play a role in allergic diseases as it causes IgE suppression [48, 49].
Based on available theoretical models, it seems that a higher TGF-β level should not have the effect found in the present study. One theory to consider is that perhaps higher TGF-β baseline levels were directly related to SARS-CoV-2 exposure in study participants, despite negative PCR results in these patients. This would explain the higher antibody production after the first dose of the vaccine. It should be remembered that the diagnosis of COVID-19 infections is not devoid of limitations, and that sensitivity of tests is often insufficient. However, other, not fully described immune mechanisms, which may correspond to an enhanced humoral response following mRNA vaccine administration, cannot be excluded. Further research is needed to explain this phenomenon.
Conclusions
This study confirmed an increase in the concentration of IgG S1 antibodies after vaccination with the mRNA preparation in all patients, regardless of gender, age and BMI. At the same time, the significantly higher concentration of IgG S1 after the first dose of the vaccine in people who have had SARS-CoV-2 infection is noteworthy.
The study did not show an effect of vaccination on an increase in levels of cytokines IL-8, IFN-γ and TGF-β, measured 3 weeks after the first dose and 3 weeks after the second dose of the vaccine. However, by an unclear mechanism, higher pre-vaccination TGF-β levels are associated with greater increases in antibody levels after the first dose of the vaccine. It is also worth paying attention to the fact that IgG N1 antibodies, indicating contact with the virus during the study, increase in some of the analysed patients and decrease in others, which indicates a significant exposure to SARS-CoV-2 infection in Poland during the study.







