
Introduction
Asthma is a common and genetically predisposed chronic inflammatory disease involving various immune-active cells and cytokines [1]. According to the World Health Organization, the global prevalence of asthma is approximately 5%. In China, the prevalence of asthma is about 4%, with regional variations ranging from 4.9% to 12.7%. Asthma is more common among populations living below the poverty line [2, 3]. Over the past few decades, the incidence of asthma in China has shown a continuous upward trend [4]. The occurrence and progression of asthma are the result of interactions between polygenic genetic factors and high-risk environmental factors [5].
Asthma is a complex genetic disease caused by a variety of genetic genes and environmental factors. A national family survey and statistical analysis have shown that patients with a family history of asthma have a significantly increased probability of asthma among their first-degree relatives, suggesting a familial genetic link to the onset of asthma [6]. Studies have identified that the 5q31-32 region on chromosome 5 plays a crucial role in regulating genetic susceptibility traits related to asthma or asthma-related phenotypes. This region contains many genes encoding immune-related molecules, including cytokine gene clusters and the recently discovered T cell immunoglobulin domain and mucin domain protein (TIM) gene family [7]. In the TIM family, TIM-3 has garnered significant attention for its role in balancing Th1/Th2 cells and regulating hypersensitivity reactions and immune responses [8]. TIM-3 is considered an inhibitory molecule for the activation of various immune cells, including Th1 cells, via providing negative regulatory signal to suppress Th1 and Tc1-mediated immune responses upon binding to its ligand. The relative reduction in Th1 cells and their cytokines, along with an enhanced Th2 immune response, are closely associated with the development of inflammation in asthma.
Current research into the polymorphism of the TIM-3 gene and its correlation with familial aggregation susceptibility to asthma has been a focus in recent years. However, the relationship’s validation in clinical practice has limitations, with no significant conclusions yet achieved. Previous epidemiological data indicated that among children with a family history of asthma, 62.2% have a family history of allergies, and 25% and 31.4% of their first- and second-degree relatives, respectively, suffer from asthma, indicating that most asthma cases have a specific genetic predisposition. If one parent has asthma, the likelihood of their children having asthma is 2 to 5 times higher than that of other healthy children. This probability increases to 10 times if both parents suffer from asthma. Identical twins have significantly higher risk of developing asthma compared to fraternal twins [9].
Aim
Our research investigated 30 asthma patients and 60 immediate family members to explore the potential relationship of TIM-3 polymorphisms with familial asthma.
Material and methods
Study subjects
This retrospective study involved patients diagnosed with asthma and their direct blood relatives who were admitted to or visited the outpatient department of Respiratory and Critical Care Medicine at our Hospital from December 2021 to December 2022. The participants were surveyed for their age, allergy history, living environment, lifestyle, gender, ethnicity, smoking history, and lung function. Data from the survey feedback, lung function, and related clinical symptoms were compiled to summarize the conditions of asthma patients and screen for high-risk factors of asthma. Based on family pedigrees, participants were divided into three direct lineage groups: Generations I, II, and III, each comprising 30 asthma patients. All subjects were free from other interfering diseases.
Inclusion criteria
Diagnostic criteria for infants with asthma in Generation III group:
1) Onset age < 3 years, with ≥ 3 episodes of wheezing; 2) Audible diffuse expiratory wheezing during acute asthma attacks, with prolonged expiratory phase; 3) Presence of atopic constitution, such as atopic dermatitis or allergic rhinitis; 4) Parental history of asthma or other allergic conditions; 5) Exclusion of other diseases causing wheezing.
A diagnosis of asthma is made if criteria 1, 2, and 5 are met. If there are 2 wheezing episodes along with criteria 2 and 5, a diagnosis of suspected asthma or wheezy bronchitis (under 3 years) is considered. If criteria 3 and (or) 4 are also present, asthma treatment may be considered.
Diagnostic criteria for children with asthma in Generation III group: 1) Age ≥ 3 years, with recurrent wheezing (if only one episode, consider if related to a specific allergen or irritant); 2) Wheezing predominantly during expiration audible in both lungs, with a prolonged expiratory phase; 3) Symptoms improve with asthma treatment or resolve spontaneously; 4) Exclusion of other diseases causing wheezing, cough, shortness of breath, or chest tightness; 5) In patients with atypical clinical presentations (e.g., no obvious wheezing or audible wheeze), at least one of the following should be present: confirmed reversible airflow limitation: a) positive bronchodilator test: inhaled rapid-acting b2-agonist results in ≥ 12% increase in FEV1 after 15 min; b) improvement in ventilatory function after anti-inflammatory treatment: administration of inhaled corticosteroids and/or anti-leukotriene medications for 4–8 weeks results in ≥ 12% increase in FEV1; c) positive bronchoprovocation test; d) maximum expiratory flow rate (PEF) daily variability (monitored over 2 weeks) ≥ 13%.
Asthma is diagnosed if criteria 1-4 or 4 and 5 are met.
Diagnostic criteria for adults with asthma in Generation I, II, and III groups: 1) Repeated occurrences of wheezing, shortness of breath, chest tightness, or cough, often related to exposure to allergens, cold air, physical irritants, chemical irritants, viral upper respiratory infections, or exercise; 2) During an episode, scattered or diffuse wheezing audible in the lungs, predominantly during the expiratory phase, with prolonged expiration; 3) Symptoms improve after treatment or resolve on their own; 4) Exclusion of other diseases causing wheezing, shortness of breath, chest tightness, and cough symptoms; 5) In patients with atypical clinical manifestations (such as no evident wheezing or physical signs), at least one positive test must be present: bronchial challenge or exercise test positive; bronchodilator test positive: FEV1 increase of 12% and an absolute increase of ≥ 200 ml; peak expiratory flow (PEF) day-night variability ≥ 10% or weekly variability ≥ 20%.
Asthma is diagnosed if criteria 1–4 or 4 and 5 are met.
Exclusion criteria
1) Exclusion of drug-specific receptor-related, pulmonary dysfunction diseases and diseases related to asthma pathogenesis: overlap with chronic obstructive pulmonary disease, obstructive sleep apnoea hypoventilation syndrome, chronic respiratory diseases due to tuberculosis, fungal infections, tumours, etc.; 2) Exclusion of asthma patients with severe cardiac, liver, kidney dysfunction, malignant tumours, hematologic, or immune system diseases; 3) Exclusion of patients with mental or legal disabilities; 4) Exclusion of subjects with poor research compliance or incomplete data.
Informed consent and ethical review
The study groups were formed based on blood relationships to construct a three-generation genetic database. All subjects voluntarily participated in this study and signed relevant agreements with researchers before the survey. The entire process of this study was monitored and ethically reviewed by the Ethics Committee of the People’s Hospital of Xinjiang Uygur Autonomous Region.
Selection of SNP sites in asthma susceptibility genes for aggregated families
SNP sites: RCT studies up to 2022 that are related to asthma susceptibility gene polymorphisms were retrieved from databases such as CNKI and PubMed. Genes linked to familial susceptibility were summarized, and core sites were selected using the GeneCards database (www.genecards.org) and the dbSNP database (https://genome-asia.ucsc.edu). SNPs in gene promoter and exon-related areas (rs953569, rs511898, and rs9313441) were chosen to test the polymorphism of high-frequency SNP sites.
Genomic DNA extraction
A total of 4 ml of venous blood was collected from all enrolled subjects, and genomic DNA from the blood was extracted using a DNA extraction kit (Cat. #2504B, Beijing Junuo Biological Technology Co., Ltd.,) following the instructions.
Primer design sequence
The selected Rs sequence numbers were referenced from the USCS gene database, and the human genome genetic polymorphism map was constructed using HapMap-CHB to analyse the distribution of high-density SNP sites. Single-base extension and PCR amplification were performed in the corresponding chromosomal bright band region of the genome. The sequence of the primers for the three core susceptible SNP sites are shown in Table 1.
PCR amplification of core gene regions
The loci of interest were amplified using a PCR kit (Cat. #AS111-01, Transgen Biotech) following the manufacturer’s instructions. The PCR products were purified using a PCR product purification kit (Cat. #D0033, Beyotime) and were sequenced by Sangon Biotech.
Statistical analysis
Based on the survey and experimental results, the acquired data on basic demographics, gene phenotypes, and allele frequencies were organized and analysed using the statistical software SPSS version 26.0. For quantitative data such as age, body mass index (BMI), and asthma history among the three groups, a homogeneity of variance test was first conducted followed by variance analysis (ANOVA).
For clinical data such as gender, ethnicity, smoking history, and allergy history, the chi-square (c2) test was used. Frequency descriptions or percentages were used to describe categorical variables for the analysis of allele frequencies and the comparison of genetic mutation patterns among different asthma groups.
For SNP loci genotype distribution, the population genetics Hardy-Weinberg equilibrium test (p2 + 2pq + q2 = 1) was applied. Comparisons of corresponding genotypes between groups were performed using Fisher’s exact test or the c2 test, as appropriate. Statistical significance was expressed using a threshold of p < 0.05, indicating that differences observed were considered statistically significant.
Results
Clinical characteristics
The comparison of the basic data among the three groups showed significant differences in age, duration of asthma history, and smoking history between the ancestral asthma group and the two control groups (p < 0.05). However, there were no statistically significant differences among the three groups in terms of gender, ethnicity, and allergy history (p > 0.05) (Table 2).
Table 2
Demographic characteristics of the studied groups
Genotyping of TIM-3 at loci rs9313441, rs511898, and rs953569
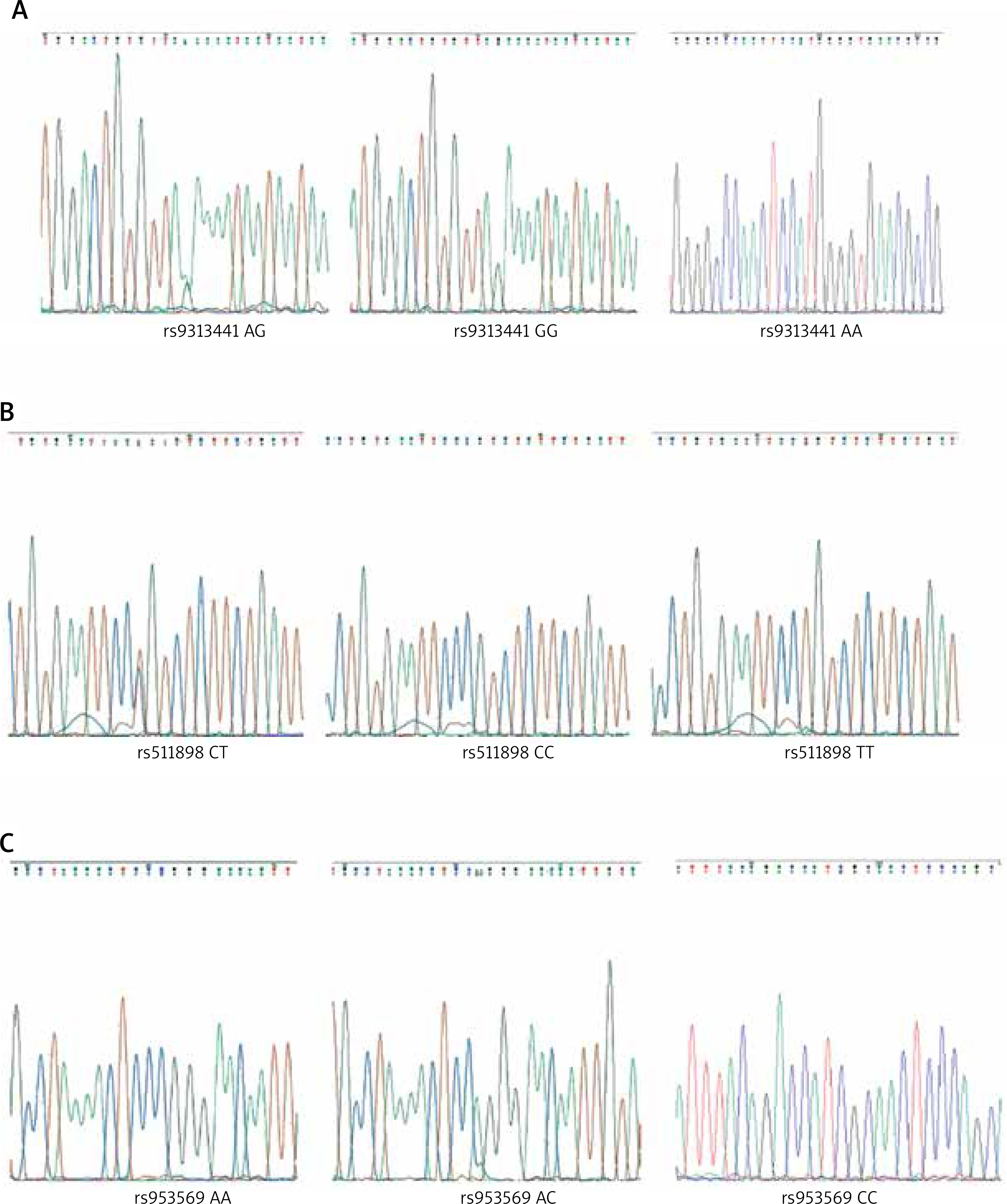
The analysis included TIM-3 family samples for SNP detection at loci rs9313441, rs511898, and rs953569, all of which exhibited gene polymorphisms in the asthma family cases studied. For the rs9313441 locus, there were three genotypes observed (GG, AG, and AA) (Figure 1 A). The rs511898 locus also presented three genotypes (CT, TT, and CC) (Figure 1 B). Similarly, the rs953569 locus showed three genotypes (CC, AC, and AA) (Figure 1 C).
Figure 1
Genotyping of TIM2 polymorphisms. A – rs9313441 polymorphism. B – rs511898 polymorphism. C – rs953569 polymorphism
Based on the gene polymorphism sequencing results, the genotyping and allele frequency expression of the TIM-3 gene loci rs953569 (C/A), rs9313441 (G/A), and rs511898 (C/T) were determined in three family groups of 30 asthma cases as follows (Table 3).
Table 3
Hardy-Weinberg equilibrium test of genotype distribution of rs953569, rs9313441 and rs511898 sites among asthmatic families (n = 30)
For rs953569, the genotype distribution frequencies of CC, AC, AA in the Generation III group were 6 (20.0%), 18 (60.0%), 6 (20.0%), respectively, with the heterozygous AC genotype being the most common. The allele frequencies were 50.0% for A and C. In the Generation II group, the genotype distribution frequencies of CC, CA, AA were 8 (26.6%), 21 (70.0%), 1 (3.34%), respectively, with allele frequencies for C and A of 38.3% and 61.6%. In the Generation I group, the genotype distribution frequencies at rs953569 for CC, CA, AA were 8 (26.6%), 21 (70.0%), 1 (3.34%), with allele frequencies for C and A of 38.3% and 61.6% (Table 3 and Figure 2 A). The analysis of gene polymorphisms for the TIM-3 gene loci rs953569 (C/A) within 30 asthma family cases revealed significant differences in the frequencies of the AA genotype at rs953569 (p = 0.032) across the three groups.
Figure 2
Hardy-Weinberg equilibrium plot and pie chart of gene frequency distribution. A – rs953569 locus. B – rs9313441 locus C – rs511898 locus
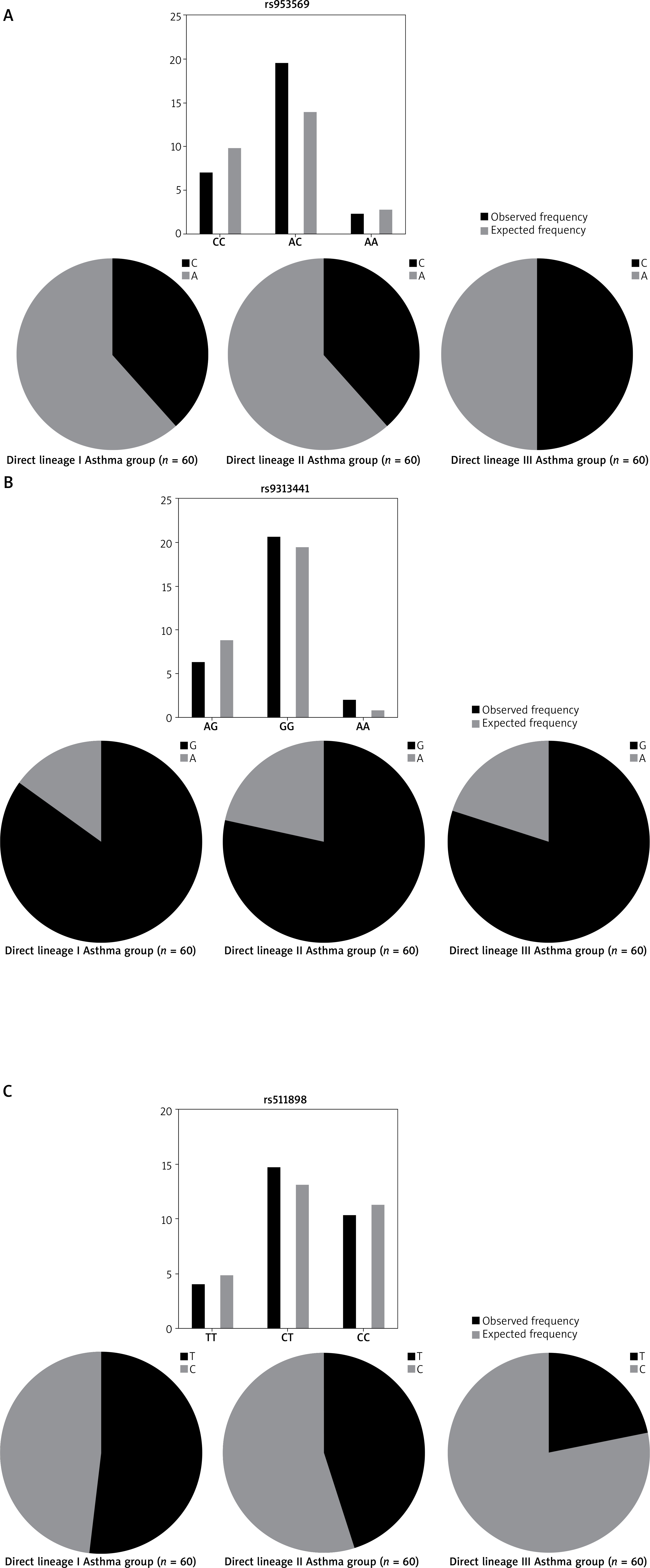
At the rs9313441 locus, the genotype distribution frequencies for GG, AG, AA in the Generation III group were 18 (60%), 12 (40%), 0 (0%), respectively, with a relatively higher expression of the GG genotype. The allele frequencies of G and A were 80% and 20%, respectively, while the distribution frequencies for GG, AG, AA in the Generation II group were 7 (23.3%), 20 (66.7%), 3 (10.0%), respectively, with allele frequencies for G and A of 78.3% and 21.7%. In the Generation I group, the genotype distribution frequencies for GG, AG, AA were 25 (83.3%), 1 (3.34%), 4 (13.3%), respectively, with allele frequencies for G and A of 85% and 15%. Statistical analysis of observed and predicted values for different genotypes showed significant differences at rs9313441 AG homozygous (p = 0.003).
For rs511898, the genotype distribution frequencies of CC, TT, CT in the Generation III group were 17 (56.7%), 13 (43.3%), 0 (0%), respectively, with the CC homozygous genotype having the highest frequency of expression. The allele frequencies of T and C were 21.7% and 78.3%, respectively. In the Generation II group, the genotype distribution frequencies of CC, CT, TT were 10 (33.3%), 13 (43.3%), 7 (23.3%), respectively, with allele frequencies for C and T of 55.0% and 45.0%. In the Generation I group, the genotype distribution frequencies of CC, CT, TT were 5 (16.6%), 19 (63.4%), 6 (20.0%), respectively, with allele frequencies for C and T of 48.3% and 51.7% (Table 3 and Figure 2 B).
Statistical analysis of observed and predicted values for different genotypes showed significant differences at rs511898 TT genotypes among the three groups (p = 0.031) (Table 3 and Figure 2 C).
Discussion
Asthma affects 300 million people worldwide, with the global incidence of asthma increasing by approximately 6–9% annually predominantly characterized by familial clustering [10]. The phenomenon of familial clustering of asthma is typically the result of gene-environment interactions, where gene-environment mediated epigenetic regulation is a high-risk factor for susceptible individuals, especially during foetal and early infant life [11–13].
Genetic susceptibility plays a significant role in the onset and progression of asthma. Using whole-genome linkage and candidate gene association approaches, over 170 genes have been identified as related to various asthma phenotypes across different populations [14]. Family studies and related epidemiological research primarily focus on the family history of close relatives of paediatric asthma patients, aiming to provide evidence for familial genetic origins and the diagnosis of specific clinical symptoms of asthma [15]. Compared to children without a family history of asthma, those with a direct relative who has asthma are significantly more likely to develop the condition. This increased risk of asthma can persist into adolescence and early adulthood. Even if neither parent has asthma, having grandparents with asthma can increase the likelihood of disease in family members [16, 17].
Members of the TIM family are encoded by three genes (HAVCR1, HAVCR2, and TIMD4) in humans, representing TIM-1, TIM-3, and TIM-4, located on chromosome regions 5q33.2 and 11B1.1, respectively. The TIM family provides a functional repertoire for the recognition of apoptotic cells, characterized by highly conserved structures and immune functions [18]. It has been confirmed that activation of the TIM-3/Gal-9 pathway can promote immune tolerance by suppressing the activity of Th1 cells, thereby positively regulating allergic diseases such as asthma and playing a key role in inflammation regulation [19]. Although there is an ongoing debate over whether TIM-3 acts as an inhibitory or stimulatory receptor in the regulation of T cell proliferation, in the pathogenesis of asthma, TIM-3 predominantly acts as a negative regulator on Th1 cells [20]. It is well known that asthma is a clinical syndrome characterized by an imbalance between Th1 and Th2 cells. Recent reports suggest that genetic polymorphisms may be the crucial factor disrupting the balance between Th1 and Th2 cells [21]. In homologous mouse models, the asthma-related TIM-3 gene has been shown to cause dysregulation of Th1 immune responses and the production of pro-inflammatory cytokines such as mouse interferon-g (IFN-g) and tumor necrosis factor-a (TNF-a) [22]. In the presence of lipopolysaccharide, stimulation of mature and functional spleen dendritic cells in mice with galectin-9 led to a significant increase in inflammatory markers such as TNF-a in mouse peripheral blood, indicating that TIM-3 regulated the induction of inflammatory transcription factor cascades [23]. Additionally, cross-linking TIM-3 on mouse mast cells, following IgE and antigen stimulation, resulted in upregulated expression and enhanced secretion of antigen-dependent Th2 cytokines IL-4, IL-6, and IL-13 [24]. These data indicate that the dynamic expression of TIM-3 in inflammatory cells is closely associated with the degree of airway inflammation, and high expression of such molecular genes may lead to a state of tolerance or incompetence to various antigenic stimuli. Activation of the TIM-3 signalling pathway by external factors may enhance inflammatory responses by inhibiting Th1 cytokine responses, upregulating Th2 cytokine responses, and inducing regulatory T cells.
Previous studies primarily focused on the relationship between TIM-3 gene polymorphisms and autoimmune and allergic diseases, such as rheumatoid arthritis, allergic asthma, and allergic rhinitis. These studies identified that certain loci of the TIM-3 gene are associated with susceptibility to these diseases, with loci such as rs1015746 and rs1036199 being strongly correlated with susceptibility to rheumatoid arthritis, and the allelic frequency of rs1015746 showing significant differences between patients with asthma and allergic rhinitis compared to control groups [25, 26]. Similarly, certain SNPs of TIM-3 have been found to be strongly associated with allergies and eczema [27]. However, genetic background variations exist between different regions and populations, such as the polymorphisms of the TIM-3 gene loci in the population of Xinjiang region not being associated with susceptibility to allergic rhinitis [28]. In the Hubei population, polymorphic variations at certain TIM-3 gene loci may be related to susceptibility to adult atopic asthma [29]. Family-based studies by Gao et al. [30] and Page et al. [31] found significant correlations between mutations in the TIM-1 gene and susceptibility to asthma and paediatric atopic dermatitis in African American and Caucasian populations. However, the polymorphisms in the TIM-3 gene were not associated with susceptibility to allergic diseases in these two populations. This study initially divided the subjects into three groups based on three generations of genetic linkage (blood relations). According to the distribution of single nucleotide polymorphisms (SNPs) from the 5′ promoter to the 3′ end of the TIM-3 gene as presented in the NCBI and UCSC gene databases, three high-frequency SNPs were selected: rs953569, rs9313441, and rs511898. These were analysed using unilateral direct sequencing. The polymorphism and allelic frequency distribution of these three genotypes were validated using Hardy-Weinberg equilibrium principles.
This study found that the CC and AC genotypes at the rs953569 locus conform to Hardy-Weinberg genetic equilibrium stability. Thus, these two SNPs exhibit genetic stability in families with asthma, ruling out their mediation of genetic susceptibility, while the homozygous AA genotype showed a significant deviation from equilibrium across the three groups. According to the genotype expression analysis at the rs953569 locus across global populations by NCBI, the AA genotype, being a recessive homozygote, could potentially represent a high-risk mutation mediating genetic susceptibility in asthma families. For the rs9313441 locus, the GG and AA genotypes are stably inherited. The heterozygous AG, due to its recombinant nature, does not allow for a definitive determination of whether the genetic gene is induced by mutation; for the rs511898 locus, the heterozygous CT genotype showed no statistically significant differences, and the CC genotype at this SNP is a dominant homozygote. However, due to the small sample size and the potential for random errors and confounding factors, the genetic stability of this genotype cannot be conclusively ruled out, and further work should increase the sample size to verify this definitively. The TT genotype frequency exhibited significant differences across the three groups, lacking genetic equilibrium stability. Further analysis by NCBI suggests that this genotype is a recessive homozygote, strongly suggesting that the TT genotype at the rs511898 locus may be a genetically susceptible gene type.
The signalling pathways of the TIM-3 subgroup can negatively regulate the activation of Th1 immune cells. However, the downstream signalling pathways of TIM-3 remain unclear in different environmental and genetic susceptibility regulatory loops. Therefore, future research should focus on identifying the activating proteins and ligand-receptor cross-linking pathways of TIM-3 molecules, which will contribute new insights into clinical interventions and the implementation of personalized treatment strategies for patients susceptible to asthma.







