
Introduction
Primary liver cancer remains a major global health challenge, with hepatocellular carcinoma (HCC) accounting for the majority of cases and ranking among the leading causes of cancer-related mortality worldwide [1]. Incidence and outcomes vary substantially across regions, reflecting differences in the distribution of risk factors as well as disparities in surveillance and access to treatment. Most cases are diagnosed in Asia, typically between 30 and 60 years of age, with male predominance. Despite improvements in prevention and therapy, the overall 5-year survival remains below 20%, underscoring the urgent need for more effective interventions [2, 3].
HCC develops through a complex interplay of non-infectious and infectious factors. Non-infectious drivers include cirrhosis, chronic alcohol intake, tobacco smoking, obesity, and type 2 diabetes. Among infectious causes, persistent infection with hepatitis B virus (HBV), hepatitis C virus (HCV), or HBV-hepatitis D virus (HDV) co-infection represents a major etiological contributor [4]. In Asia, an epidemiological shift has been noted from viral hepatitis-driven HCC to non-alcoholic fatty liver disease (NAFLD) and its progressive form, non-alcoholic steatohepatitis (NASH). Cirrhosis, whether viral or metabolic in origin, remains one of the strongest risk factors for malignant transformation; however, HCC can also arise in NAFLD without cirrhosis. Notably, the coexistence of HBV infection and fatty liver in Asian populations accelerates fibrosis progression and HCC development [5].
Additional exposures further influence disease burden. In Africa and Southeast Asia, the dietary ingestion of aflatoxin B1, produced by Aspergillus species, synergistically enhances the risk of HCC in individuals with HBV infection. The recently defined entity metabolic dysfunction-associated steatotic liver disease (MASLD) also contributes to hepatocarcinogenesis by promoting fat accumulation, inflammation, and immune dysregulation within the hepatic microenvironment. While early-stage tumours are potentially curable, most cases are diagnosed late; hence, risk-based surveillance remains a cornerstone of improving survival [6].
Parallel to advances in systemic therapies, there is increasing interest in natural compounds with anticancer properties. Campesterol, a dietary phytosterol abundant in nuts, seeds, cereals, and vegetable oils, is synthesised via the mevalonate pathway and shares structural similarity with cholesterol. Traditionally recognised for its cholesterol-lowering, anti-inflammatory, and antimicrobial effects, campesterol has recently been implicated in cancer prevention. Experimental studies suggest that it exerts anti-proliferative and pro-apoptotic effects by modulating oxidative stress, reactive oxygen species, antioxidant enzyme activity, and apoptotic pathways [7].
Based on this background, we hypothesised that campesterol would decrease viability and promote apoptosis in HepG2 liver cancer cells. The present study, therefore, aimed to evaluate the effect of campesterol on cell proliferation, viability, and apoptosis in vitro, while also defining its dose-response relationship.
Material and methods
Cell culture
The human hepatocellular carcinoma cell line HepG2 was obtained from the cell culture facility at the University of Lahore. Cells were preserved in liquid nitrogen until required. For recovery, cryovials were thawed rapidly at 37°C and transferred into culture flasks containing Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1× penicillin-streptomycin. Cultures were maintained at 37°C in a humidified atmosphere with 5% CO2. Subculturing was performed at 70-80% confluence using 0.25% trypsin-EDTA, as previously described [8].
Preparation of campesterol
Campesterol was obtained from the Department of Pharmacy at the University of Lahore. Stock solutions were prepared in culture medium and diluted to final concentrations of 10, 50, 100, and 200 μg/ml for use in the experiment.
Growth medium preparation and MTT assay
DMEM was supplemented with 10% heat-inactivated FBS and 1× penicillin-streptomycin solution. The complete medium was sterilised by filtration through a 0.2 μm membrane filter and stored at 4°C until use. Cell viability was assessed using the MTT assay [9]. Briefly, HepG2 cells were seeded into 96-well plates and treated with campesterol (10, 50, 100, and 200 μg/ml) for 24 h. After treatment, wells were washed with phosphate-buffered saline (PBS) and incubated with 100 μl of serum-free medium containing 25 μl of MTT solution (0.5 mg/ml) for 3 hours at 37°C. Formazan crystals were solubilised with 10% sodium dodecyl sulphate (SDS), and absorbance was measured at 570 nm using a microplate reader. Viability was calculated relative to untreated controls.
Crystal violet staining assay
Cell viability was also evaluated using crystal violet staining. HepG2 cells were seeded in 96-well plates and treated with campesterol as above. Following treatment, the cells were washed with PBS, fixed with 2% ethanol, and stained with a 0.1% crystal violet solution for 15 minutes at room temperature. Wells were washed thoroughly to remove excess dye, and the bound stain was solubilised with 100 μl of 1% SDS for 5-10 min. Absorbance was recorded at 540 or 595 nm using a microplate reader.
Statistical analysis
All experiments were performed in triplicate and repeated at least three times independently. Data are presented as mean ± standard deviation (SD). Statistical comparisons between groups were carried out using one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test to identify pairwise differences. A p-value of < 0.05 was considered statistically significant. Analyses were performed using GraphPad Prism (version X.X, GraphPad Software, San Diego, CA, USA).
Results
Effect of campesterol on HepG2 cell viability (MTT assay)
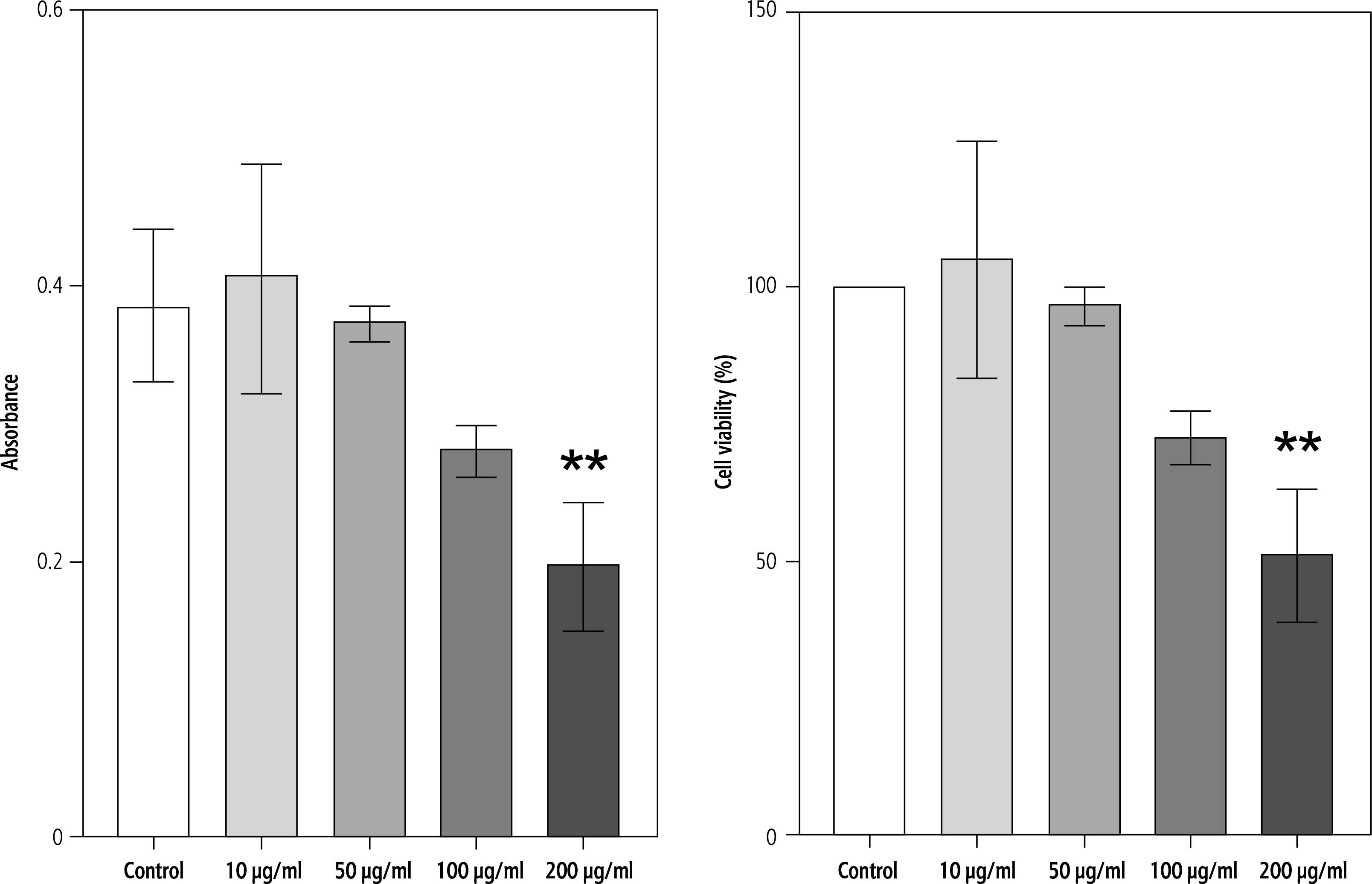
The effect of campesterol on HepG2 cell viability was evaluated using the MTT assay after 24 h of treatment. Cells were exposed to increasing concentrations (10, 50, 100, and 200 μg/ml). Campesterol did not significantly alter viability at lower concentrations (10-100 μg/ml). However, a marked reduction in cell viability was observed at 200 μg/ml compared with untreated controls (p < 0.05, one-way ANOVA followed by Tukey’s post-hoc test) (Fig. 1).
Fig. 1
Cytotoxicity rate of HepG2 cells after treatment with campesterol extract by MTT assay. Effect of campesterol on HepG2 cell viability measured by MTT assay. Data are presented as mean ± SD from three independent experiments. Significantly lower cell viability was observed at 200 μg/ml compared with the control (*p < 0.05)
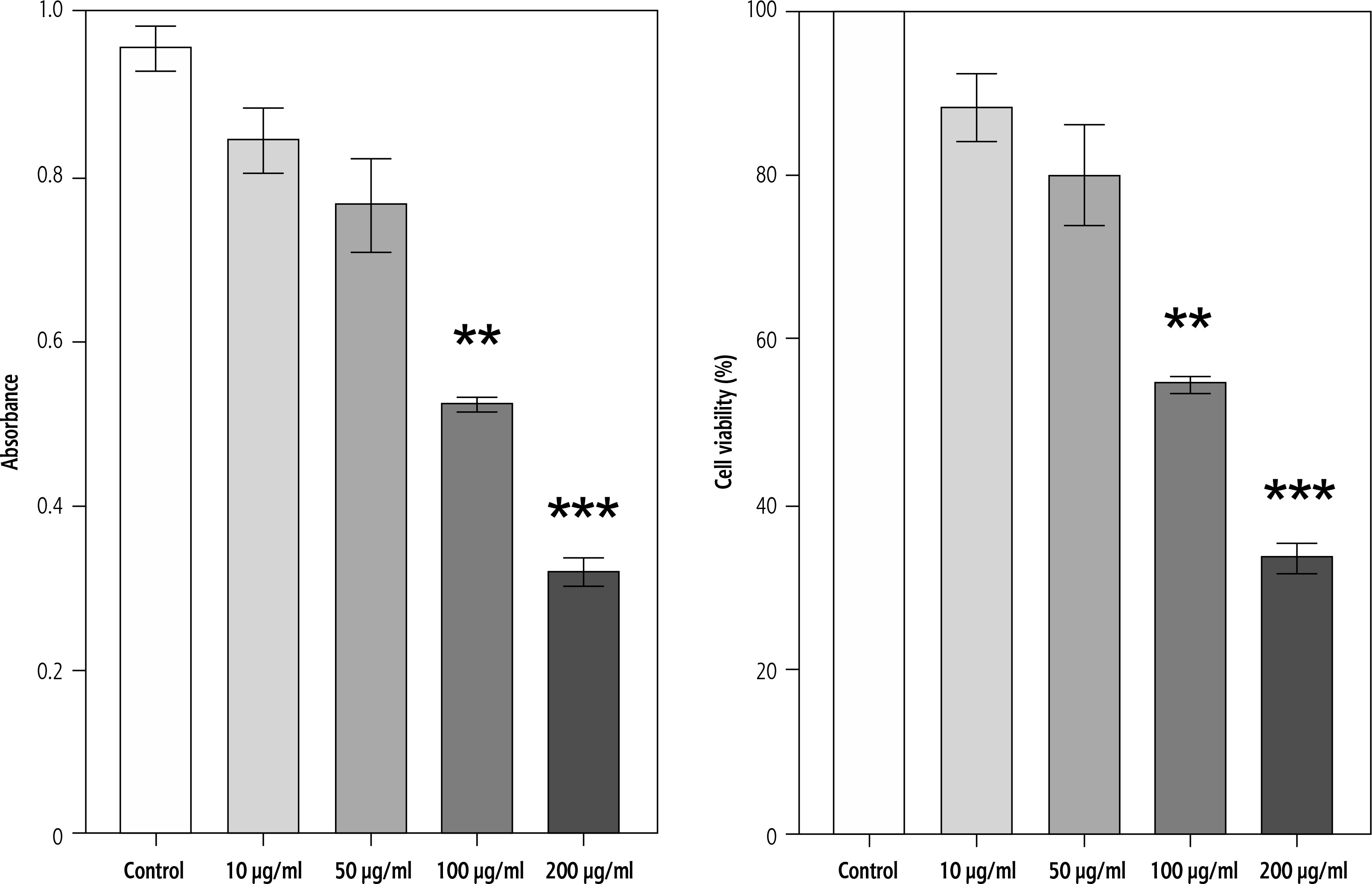
Effect of campesterol on adherent cell biomass (crystal violet assay)
To further confirm the effect of campesterol on cell viability, crystal violet staining was performed. Similar to MTT results, HepG2 cells treated with campesterol at 200 μg/ml showed significantly reduced staining intensity compared with untreated controls, indicating a reduction in adherent viable cells. No significant differences were detected at lower concentrations (10-100 μg/ml) (Fig. 2).
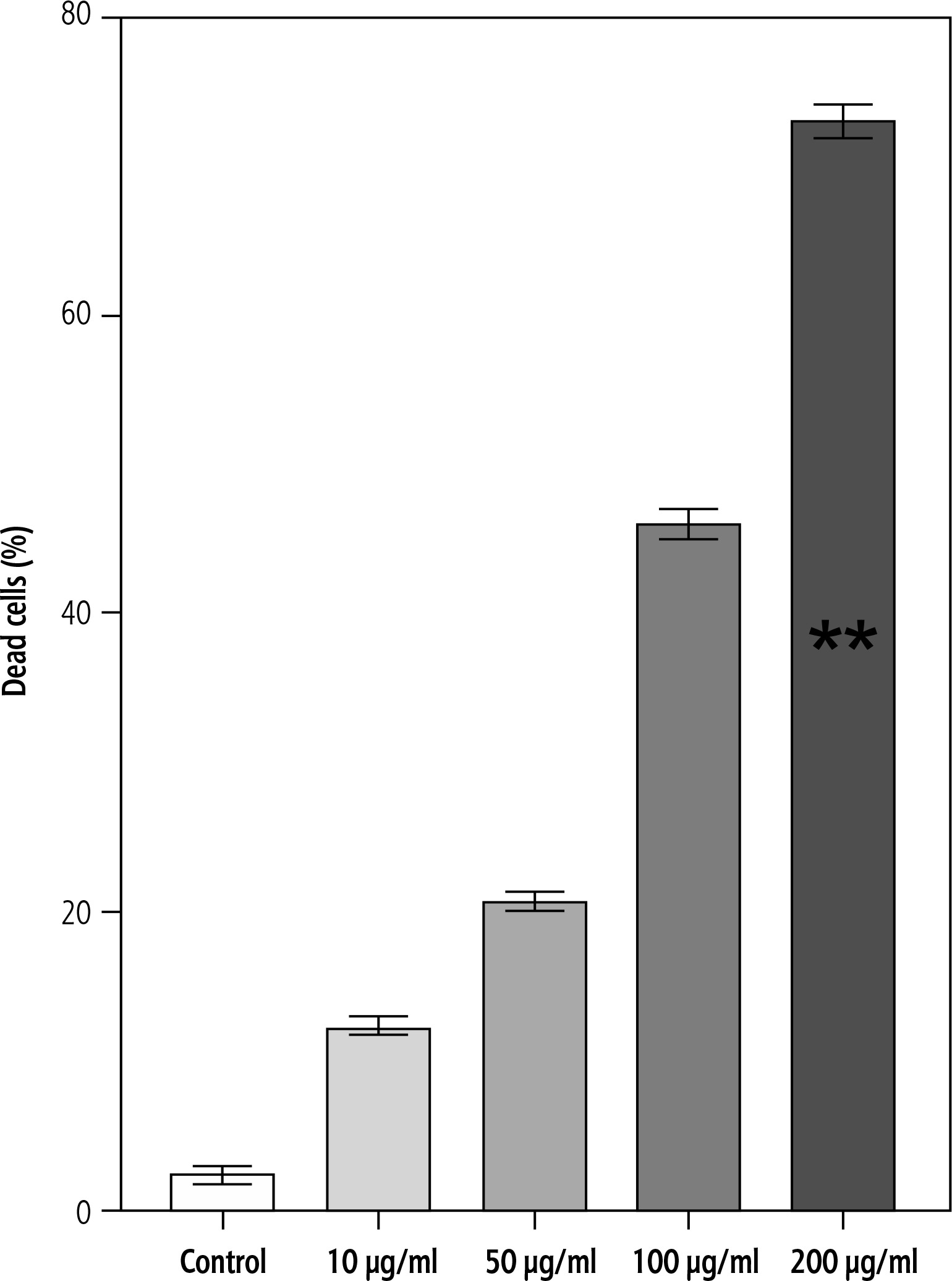
Effect of campesterol on cell death (trypan blue assay)
Cell death was further quantified using trypan blue exclusion. Exposure of HepG2 cells to campesterol for 24 h induced a concentration-dependent increase in the proportion of non-viable cells. The most pronounced effect was observed at 200 μg/ml, where the percentage of dead cells was significantly higher than in controls (p < 0.05) (Fig. 3).
Discussion
Liver cancer remains one of the most lethal malignancies, with HCC representing 75-85% of cases and intrahepatic cholangiocarcinoma comprising the majority of the remainder [10, 11]. Despite preventive strategies and therapeutic advances, the global incidence of HCC continues to rise, and prognosis remains poor. This underscores the need for alternative or adjunctive therapeutic approaches, including the exploration of natural bioactive compounds with anticancer potential [12-14]. Over the past decades, natural products have emerged as a rich source of anticancer agents. Many plant-derived compounds exert cytotoxic effects by modulating key signalling pathways involved in cell proliferation, apoptosis, and metastasis [15, 16]. Among these, phytosterols have garnered increasing attention due to their favourable safety profile and broad range of biological activities, including anti-inflammatory, cholesterol-lowering, and anticancer effects [17-20].
In this study, campesterol, a dietary phytosterol, was found to reduce viability and promote apoptosis in HepG2 liver cancer cells. Across three complementary assays – MTT, crystal violet, and trypan blue – campesterol consistently decreased proliferation and increased cell death at higher concentrations (200 μg/ml) [21, 22]. These findings are in agreement with previous studies, which have shown that sterol derivatives, such as 7-ketositosterol and β-sitosterol [23], suppress proliferation and induce apoptosis in liver, breast, and ovarian cancer cells through mechanisms involving p53 signalling [24], mitochondrial dysfunction, and endoplasmic reticulum stress [25, 26].
Apoptosis is a central target in anticancer drug development, as tumour cells often acquire the ability to evade programmed cell death. Our results suggest that campesterol triggers apoptotic pathways in HepG2 cells, as evidenced by reduced viability and increased non-viable cell fractions. Although our assays confirmed cytotoxicity, further mechanistic studies, such as Annexin V/PI staining, caspase-3/7 activation, PARP cleavage, and mitochondrial membrane potential assays, are necessary to establish the molecular events underlying campesterol-induced apoptosis conclusively.
An essential strength of this study is the use of multiple viability assays to validate the cytotoxic effect of campesterol. However, there are limitations. First, only one liver cancer cell line (HepG2) was tested; additional HCC models are required to assess generalizability. Second, mechanistic apoptosis assays were not performed, which limited our ability to pinpoint the precise signalling pathways involved. Finally, this was an in vitro study; in vivo validation and pharmacokinetic profiling are essential before clinical translation can be considered.
Despite these limitations, our findings support the growing evidence that dietary phytosterols possess anticancer potential. Campesterol, in particular, may represent a promising low-toxicity adjunct for liver cancer management. Future studies should expand to multiple liver cancer models, explore synergistic interactions with established agents such as sorafenib or lenvatinib, and investigate the translational potential of phytosterols in vivo.
Conclusions
This study demonstrates that campesterol, a dietary phytosterol, reduces cell viability and promotes apoptosis in HepG2 liver cancer cells in a dose-dependent manner. By integrating complementary viability assays, our findings provide preliminary evidence for the anticancer potential of campesterol. While the precise molecular mechanisms remain to be elucidated, these results highlight phytosterols as promising candidates for low-toxicity adjuncts in the treatment of hepatocellular carcinoma. Future studies should extend to additional liver cancer models, incorporate mechanistic apoptosis assays, and evaluate in vivo efficacy to determine the translational applicability of campesterol in cancer management.
Disclosures
This research received no external funding.
This study was approved by the biomedical ethical committee (Ref-IMBB-BBBC-24-00-C). The study was conducted according to the guidelines of the Declaration of Helsinki. All participants provided informed consent.
The authors declare no conflict of interest.





