
Introduction
During the past decade, the impact of the expression of corticotropin-releasing hormone (CRH) family peptides and their corresponding receptors on human physiology and onset of disease has gained the attention of the scientific community. However, the complex interplay of the numerous biological processes often leads to controversial study results. It seems that, despite the significant progress made in understanding stress pathophysiology and its implications over the past few years, we are still only scratching the surface of the apparent relationship between stress family peptides, inflammation, and carcinogenesis. All living organisms – including humans – are forced to respond to numerus stimuli daily which act as stressors. The stress response system is designed to promote survival through homeostasis alteration and activation of complex neurobiological systems, as well as behavioural modifications, which aim to eventually reach a new equilibrium state, or allostasis. However, when frequent neurobiological response to stressors is needed or intense stress arises, the risk of onset of illnesses increases [1]. This new deregulated state of the stress response system – either in terms of duration or intensity – is known as “cacostasis” and has adverse effects on a wide range of biological functions [2].
Corticotropin-releasing hormone and related peptides (urocortins 1–3), in adjunct with their related receptors CRHR1 and CRHR2, are major regulators of the human stress response axis [3]. Nevertheless, their expression has been demonstrated not only in tissues of the central nervous system, but also in various peripheral organs, such as the skin, heart, lungs [4, 5], and most tissues of the female reproductive system [6–9]. Their peripheral biosynthesis has also been demonstrated in a large variety of human cancer cell lines, such as neuroblastomas, small-cell lung cancers, and melanomas [10–14], with their exact biological role in malignancy remaining elusive and a subject of debate among researchers.
In this study, we sought to systematically review the existing literature regarding the expression of CRH family peptides and their receptors in gynaecological malignancies and their known premalignant conditions, including endometrial, ovarian, vulvar, cervical, and breast cancer. Furthermore, we summarize current knowledge regarding their possible implications in tumourigenesis and progression of malignancy. By summarizing existing knowledge, we aim to attract the attention of the scientific community on the multifaceted roles of CRH family peptides and their possible influence on tumourigenesis and tumour progression in gynaecological cancers, providing a basis to prompt for further research.
Material and methods
To explore the expression and possible effects of CRH family peptides and their receptors on gynaecological malignancies and their precancerous lesions, we utilized the medical search engines PubMed, Scopus, and Cochrane Library. Our search strategy was the following: (“stress hormones” or “stress axis” or “CRH” or “corticotropin-releasing factor” or “urocortin” or “UCN” or “CRHR1” or “CRHR2”) and (“endometrial cancer” or “ovarian cancer” or “cervical cancer” or “vulvar cancer” or “fallopian tube cancer” or “breast cancer” or “vaginal cancer” or “gynecological cancer” or “CIN” or “VIN” or “VaIN” or “gynecological premalignant conditions” or “gynecological precancerous conditions” or “endometrial hyperplasia”). We sought human, prospective, and retrospective cohort studies, systematic reviews, and meta-analyses in English language, published in medical journals prior to 1 September 2022.
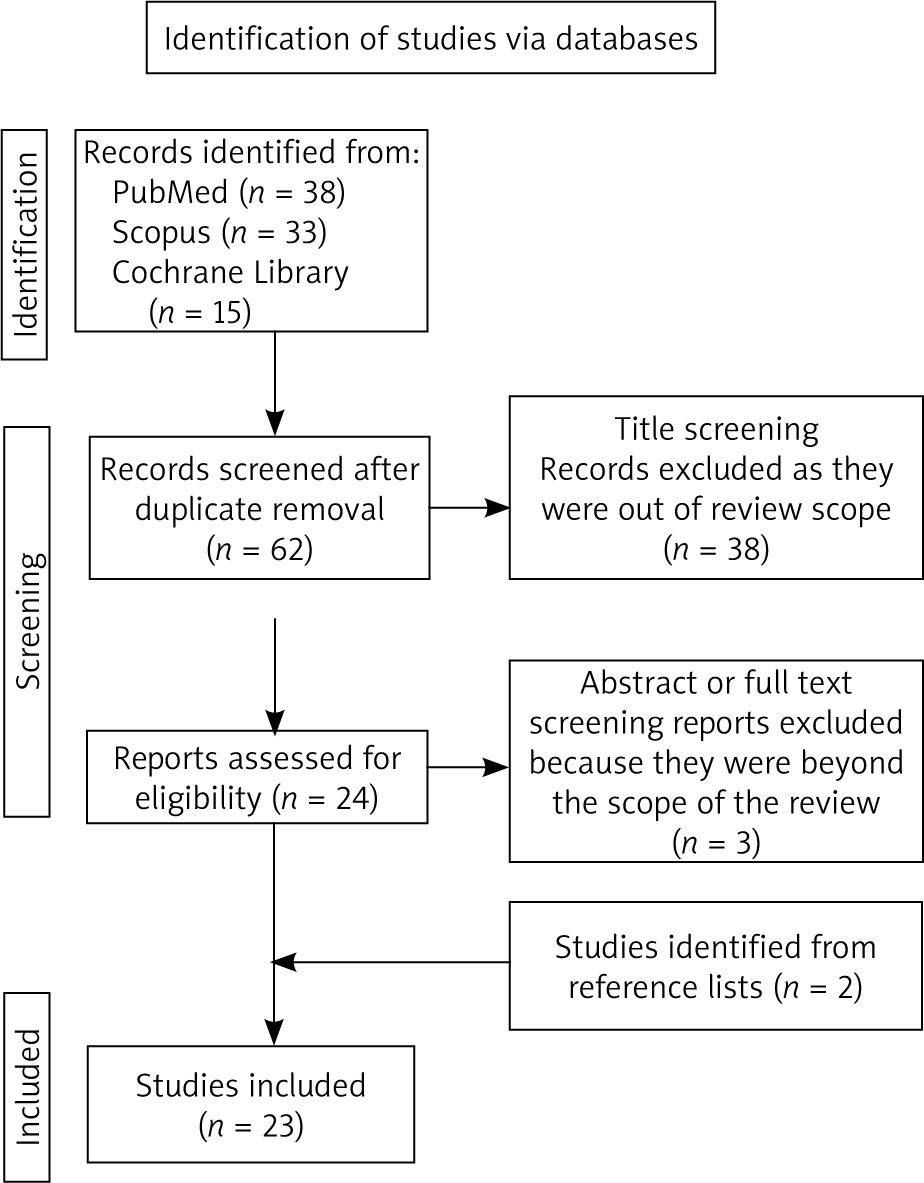
Our search initially detected 71 published papers. After removal of duplicates, 47 studies underwent the title screening phase by 2 independent reviewers. Twenty-three studies were excluded during this process because they were outside the scope of the review. The remaining 24 full-text papers were assessed for eligibility, and 21 relevant studies were selected for review. Reference lists of relevant studies were screened for studies not detected by our initial search. Finally, 23 studies were included in our systematic review. The flow chart of the study is depicted in Figure 1.
Results
Breast cancer
Our literature review yielded 10 studies that referred to the correlation of CRH family peptides and breast cancer, and these are briefly summarized in Table 1. The first attempt to detect CRH in breast cancer cells was in 1990. Ciocca et al. [15] detected CRH expression in 30% of breast cancer biopsies (colloid, lobular, and infiltrating ductal carcinomas) but found no clear relationship between CRH staining and tumour grade or stage. Nevertheless, CRH staining was observed in nearly 20% of cases with atypical hyperplasia, while it was absent in normal mammary cells. In his preliminary study, Graziani et al. [16] validated the autocrine and/or paracrine effects of CRH on tumoural cell proliferation and/or differentiation, suggested by Ciocca et al., in MCF7 breast cancer cells.
Table 1
Studies detected in literature concerning the expression and possible implications of corticotropin-releasing hormone family peptides in human breast cancer
| Author | N | Type of study | Keypoint results |
|---|---|---|---|
| Ciocca et al. [15] | 40 | Retrospective study | CRH expression detected in 30% of breast cancer biopsies. No clear relationship of CRH expression status with patent’s clinicopathological indices |
| Graziani et al. [16] | N/A | In vitro study | MCF7 cells express CRH mRNA and secrete sizable amounts of immunoreactive CRH. Validation of autocrine and/or paracrine effects of CRH on MCF7 breast cancer cells |
| Kaprara et al. [17] | 29 | Retrospective clinical study | CRHR1 and CRHR2 gene transcript expression in 23% and 36% of breast cancer cases, respectively |
| Koureta et al. [18] | N/A | In vitro study | of corticotropin-releasing factor (CRF)-R1 and CRF-R2 is up- and down-regulated, respectively, by exposure to oestradiol activation of CRF-R2 increased the migration of MCF-7 cells and potentiated an oestrogen-inducing effect |
| Androulidaki et al. [19] | N/A | In vitro study | CRH inhibits apoptosis and proliferation but enhances motility and invasiveness of MCF-7 cells via CRHR1 activation |
| Jin et al. [21] | N/A | In vitro study | CRH and urocortin 2 induce concentration-dependent apoptosis of MCF-7 cells |
| Jin et al. [22] | N/A | In vitro study | CRH suppresses TGFβ1-induced EMT and upregulates e-cadherin expression |
| Jin et al. [23] | N/A | In vitro study | Urocortin inhibits TGFβ1-induced Snail1 and Slug mRNA expression, disrupts TGFβ1 signalling-mediated EMT, and attenuates Smad2/3-TGFβ1 signalling |
| Jin et al. [25] | N/A | In vitro study | CRHR1 and CRHR2 inhibit autophagosome formation and impede migration potential in MCF-7 malignant cells |
| Balogh et al. [20] | N/A | In vitro study | Urocortin treatment activates ERK1/2, increases tumour protein p53 expression, and reduces ATP production in MCF-7 cells |
In their small-sampled series of human breast cancer biopsies, Kaprara et al. [17] observed CRHR1 and CRHR2 gene transcript expression in 23% and 36% of cases, respectively. In fact, CRHR1 was detected in cancerous implants and epithelial cells of breast ducts, whereas CRHR2 was mainly noted in perineural invasions with their subcellular localization being both cytoplasmic and membranic. No significant associations with patient histopathological indices were noted. The authors concluded that the 2 receptors have distinct biological roles and may be implicated in a complex interplay between malignant cells and other proximal cells, affecting tumoural growth, metastatic potential, and/or immunosurveillance [17].
Recently, the same study group reported the in vitro detection of both CRHR1 and CRHR2 mRNAs in MCF-7 cell lines. Oestradiol exposure increased CRHR1 mRNA levels, while both CRHR2 subtypes (a and b) decreased [18]. Treatment with UCN2, a selective agonist of CRHR2, increased cancer cell migratory potential and potentiated the effects of oestradiol on migration but had no effect on proliferation [18]. These findings were consistent with those of Androulidaki et al. [19], who reported that CRH inhibits apoptosis and proliferation but enhances motility and invasiveness of MCF-7 cells via CRHR1 activation. FAK protein activation and increased COX-1 production were also observed, indicating a CRH-induced phenotype favouring metastasis. However, CRHR1b and CRHR2a receptor subtypes were not detected in MCF-7 cells [19]. Another recent study confirmed positive cytoplasmic expression of CRHR1 and CRHR2 in MCF-7 cells [20].
The effects of CRH family peptides on breast tumourigenesis remain controversial. Jin et al. [21], after establishing the ubiquitous detection of urocortin mRNA in MCF-7 cells, reported that CRH and Urocortin 2 induced concentration-dependent apoptosis of cancer cells presumably by androgen and vitamin D receptor upregulation and heat shock protein 27 activation. Contradicting previous findings, the same study group reported that CRH expression in breast cancer may favour more aggressive phenotypes. They also demonstrated that both CRHR1 and CRHR2 are expressed in MCF-7 cells, while only CRHR2 is expressed in MDA-MB-231 cells [22]. Corticotropin-releasing hormone suppressed TGFβ1-induced epithelial-mesenchymal transition (EMT) – generally considered as the first step of metastatic cascade – and upregulated e-cadherin expression via both receptors, suggesting a metastasis-inhibiting role for CRH in breast cancer [22]. In a later study, urocortin inhibited TGFβ1-induced Snail1 and Slug mRNA expression, disrupted TGFβ1 signalling-mediated EMT, and attenuated Smad2/3-TGFβ1 signalling through Smad7 upregulation [23]. Of note, Snail1 and Slug are considered negative transcript regulators of E-cadherin, and their upregulation by TGFβ1 plays a key role in EMT transformation of cancerous cells [24]. The above findings led authors to suggested that urocortin may be utilized as a novel therapeutic agent in breast malignancies disrupting EMT via attenuation of TGFβ1-induced Smad2/3 activation and Smad7 upregulation [23]. A recent study linked CRH receptor signalling to autophagy inhibition in malignancies [25]. Both CRHR1 and CRHR2 inhibited autophagosome formation, impeding LC3BI conversion to LC3BII through PLCβ-IP3-mTOR signalling pathway activation. This suggests that CRH receptor activation inhibits autophagy and hampers migration potential in MCF-7 malignant cells [20, 25]. Lastly, Balogh et al. reported that urocortin treatment of MCF-7 cells rapidly activated ERK1/2 through phosphorylation and facilitated its translocation to the nucleus. They also demonstrated increased expression of the tumour protein p53 and reduced ATP production by malignant cells after urocortin treatment, indicating increased cell proliferation and survival in MCF-7 cells. These effects could potentially be targeted with CRHR1- or CRHR2-specific inhibitors [20].
Endometrial cancer
Eight studies were identified that referred to the correlation of CRH family peptides with gynaecological malignancies (Table 2). It was as early as 1995 when Makrigiannakis et al. demonstrated the in vitro expression of immunoreactive CRH, not only in normal, non-pregnant human endometrial epithelium, but also in an established cell line of a well-differentiated human endometrial adenocarcinoma (Ishikawa cells). Using mRNA extracts of Ishikawa cells, both CRH mRNA and its peptide product were identified in cytoplasmic granules, confirming peripheral CRH expression in endometrial malignancy. Some years later, Graziani et al. [26] showed that CRH inhibited cell proliferation in Ishikawa cells via autocrine and/or paracrine mechanisms, mediated by CRHR1 activation. However, given previous reports correlating CRH-CRHR1 expression with tumour progression in melanoma cells [13], they postulated that CRH effects may be tissue-specific and dependent on receptor subtype expression. In their subsequent pilot study, however, the same authors demonstrated that CRHR1 expression was associated with more aggressive endometrial tumours [27].
Table 2
Studies detected in the literature concerning the expression and possible implications of corticotropin-releasing hormone H family peptides in human endometrial cancer
| Author | N | Type of study | Keypoint results |
|---|---|---|---|
| Makrigiannakis et al. [7] | N/A | In vitro study | Immunoreactive CRH expressed in normal endometrial epithelium and adenocarcinoma cell line |
| Graziani et al. [26] | N/A | In vitro study | CRH inhibits cell proliferation in Ishikawa cells via CRHR1 activation. CRH-R1 |
| Graziani et al. [27] | 19 | Pilot study | CRHR1 expression associated with more aggressive endometrial tumours |
| Miceli et al. [28] | 51 | Retrospective clinical study | Diffuse cytoplasmic staining of CRH, CRHR1, and CRHR2 in endometrial cancer samples. Positive correlations were found between CRH-R1 and PR expression, as well as between CRH-R2 cytoplasmic pattern, and more advanced FIGO stage disease, respectively |
| Sato et al. [29] | 87 | Retrospective clinical study | CRHR1 expression associated with worse prognosis and higher recurrence rate. CRHR1 is a potent prognostic factor |
| Jo et al. [30] | N/A | In vitro study | CRH enhances invasiveness and migration of Ishikawa cells through MMP-2 and MMP-9 upregulation |
| Florio et al. [31] | 22 (13 controls) | Retrospective clinical study | Diminished expression of UCN mRNA in endometrial carcinomas |
| Owens et al. [32] | In vitro study | UCN1 treatment suppresses migration potential of malignant cells via CRHR2 activation |
Miceli et al., using immunohistochemistry (IHC) on 51 tissue samples with confirmed endometrial cancer, reported a diffuse cytoplasmic staining of CRH, CRHR1, and CRHR2 [28]. Furthermore, higher CRHR1 expression was associated with positive progesterone receptor expression, and thus with a more differentiated and less aggressive tumoural phenotype, while higher CRHR2 expression correlated with advanced disease stage. Nevertheless, no differences in disease-free survival were observed between cases according to CRH, CRHR1, or CRHR2 status during the follow-up period of 51 months. Similarly, Sato et al. [29] confirmed these findings, reporting a worse prognosis and higher recurrence rate in cases with CRHR1 expression, while CRHR2 expression tended to correlate with a better clinical outcome. The authors concluded that CRH-CRHR1 signalling seems to play an important role in endometrial cancer progression.
Although the expression of CRH family peptides is well documented in endometrial malignancy, their exact functions are still unclear. Jo et al. [30] in their study demonstrated that treatment of Ishikawa cells with CRH significantly enhances their invasiveness and migration in vitro. The increased potential of invasiveness was suggested to be mediated by upregulation of the metalloproteinases MMP-2 and MMP-9. Following the identification of UCN expression in normal endometrial tissues, Florio et al. [31], reported a diminished expression of UCN mRNA in endometrial carcinomas compared to healthy postmenopausal samples, suggesting a reduced inhibitory effect on malignant cell proliferation.
The above suggestion was investigated by Owens et al. [32]. Using an established Ishikawa cell line, they showed that UCN1 treatment suppressed migration potential of malignant cells via CRHR2 activation. UCN1 expression was also inhibited by oestrogen receptor (ERA) activation, while oestrogen receptor (ERB) activation had no effect, in line with previous work by Makrigiannakis et al. [33], who demonstrated that oestradiol inhibits the activity of CRH promoter in human adenocarcinoma cells. Lastly, the reported downregulation of ERA and ERB expression by UCN1 led researchers to postulate that loss of UCN activity in endometrial cancer could enhance oestrogen-induced proliferation and migration of malignant cells.
In conclusion, the published literature seems to suggest that the correlation between the expression and implication of CRH family peptides and endometrial cancer is still not fully understood, with contradictory findings regarding the expression of CRHR1 and the impact of CRH on cell proliferation and migration. Further research is needed to unravel the complex mechanisms and identify potential clinical implications of CRH family peptides in this type of malignancy.
Ovarian cancer
Only 2 studies were identified in the literature regarding the expression and role of CRH and similar peptides in ovarian cancer (Table 3). Following human CRH sequencing in 1983 [34], Suda et al. showed that immunoreactive CRH was present in homogenized human ovarian cancer tissues [35]. However, it was not until 2007 that Minas et al. [36] sought to investigate the potential involvement of intratumoural CRH in this type of gynaecological malignancy, using samples of human ovarian cancer, as well as OvCa3 and A2780 ovarian cancer cell lines. In addition to CRH peptide, researchers demonstrated in vitro the expression of CRHR1, CRHR2 (not present in OvCa3 cells), as well as the proapoptotic molecule Fas ligand (or FasL), using immunohistochemical methods. The subcellular localization of CRH peptide was cytoplasmic, whereas CRHR1, CRHR2, and FasL were mainly detected on the cell membranes. Moreover, increased CRH and FasL staining correlated with increased tumoural stage. Further research in ovarian cancer cell lines revealed that incubation with CRH promotes CRHR1-mediated FasL overexpression but has no effect on tumoural cell growth.
Table 3
Studies detected in literature concerning the expression and possible implications of corticotrophin-releasing hormone family peptides in gynaecological malignancies (ovarian, cervical, and vulvar cancer)
| Author | Type of malignancy | N | Type of study | Keypoint results |
|---|---|---|---|---|
| Suda et al. [35] | Ovarian | 2 | In vitro study | Immunoreactive CRH present in human ovarian cancer tissues |
| Minas et al. [36] | Ovarian | 47 | Retrospective | CRH, CRHR1, CRHR2, and FasL were expressed in 68.1, 70.2, 63.8, and 63.8% of the cases, respectively. Positivity for CRH or FasL expression was associated with higher tumour stage |
| Taliouri et al. [39] | Cervical | N/A | In vitro study | CRH peptide expression and FasL induction by CRH in human cervical cancer cells |
| Song et al. [40] | Cervical | N/A | In vitro study | Positive CRH and CRHR2 gene expression, ULBP2 upregulation, and NKG2D receptor downregulation in HeLa cells after CRH treatment |
| Dimas et al. [41] | Vulvar | 87 | Retrospective | Increasing cytoplasmic signal of CRH, urocortin, Fas and FasL from precancerous lesions to VSCC samples. Nuclear localization of UCN in both premalignant and VSCC lesions, with staining being significantly intensified in carcinomas, particularly in the less differentiated tumour areas or in the areas at invasive tumour front |
Fas ligand is a molecule released from activated T and NK cells. Its binding to its specific receptor Fas induces apoptosis to Fas-bearing cells [37]. Based on this, researchers in the same study investigated the effect of CRH-exposed ovarian cancer cells on activated lymphocytes. The results suggested that there was a significant induction of activated lymphocyte apoptosis possibly mediated by Fas/FasL apoptotic compound [36]. Similar results regarding the autocrine/paracrine actions of intratumoural CRH have also been reported previously in both animal and human cell lines [37, 38].
Cervical cancer
Similarly, 2 studies were also identified regarding CRH family peptides in cervical malignancies (Table 3). Taliouri et al. [39] were the first study group that demonstrated CRH peptide expression with cytoplasmic localization in human cervical cancer cells belonging to the HeLa cervical carcinoma cell line. They also reported a positive mRNA expression of both CRH and FasL, as well as a FasL expression induction by CRH, in line with previous results in ovarian malignancies [36]. The authors concluded that CRH tumoural expression may contribute to a local tumoral immunoescape via FasL expression upregulation. Similarly, Song et al. [40] reported a strong positive CRH and CRHR2 – but not CRHR1 – gene expression in HeLa cells. Additionally, after CRH treatment of HeLa cells, the expression of the soluble form of ULBP2 – a ligand for NKG2D receptor on the surface of natural killer cells (NK) – was found to be significantly upregulated in malignant cells in a dose-dependent manner. Contrarily, surface expression of ULBP2 was unaffected. Further research revealed that CRH expression by HeLa cells can potentially inhibit NK cell function through NKG2D receptor. The authors suggested that CRH produced by cervical cancer cells may potentiate the evasion of the host’s cell-mediated immune response by malignant cells via ULBP2 overexpression and thus NKG2D receptor downregulation in NK cells.
Despite these promising results, the expression of CRH and related peptides as well as their possible role in tumourigenesis have not been investigated using cervical cancer tissue samples. If immunoescape of malignant cervical cells through either FasL or NKG2D or both is verified in vivo, new anti-CRH therapeutic strategies may be an option to restrict tumour progression in cervical cancer.
Vulvar cancer
Regarding vulvar malignancies, only one study was identified (Table 3). To our knowledge, our research team is the only one having already investigated the presence and implications of CRH family peptides in both human vulvar premalignant conditions and vulvar cancer samples using IHC to date [41, 42].
We observed an increasing cytoplasmic signal of CRH and urocortin from precancerous lesions (VIN and lichen) to vulvar squamous cell carcinoma (VSCC) samples. Furthermore, urocortin was also localized in the nucleus of VSCC and premalignant lesions, with intensified staining in carcinomas, particularly in less differentiated malignant cells and in those belonging to invasive tumour front, suggesting a role in tumour progression. CRHR1 expression was scarce in VSCC samples compared to premalignant lesions but of statistical significance. CRHR2 expression was weak in all conditions studied. We also investigated the possibility of vulvar cancer escaping immunosurveillance through the Fas/FasL signalling pathway. Fas expression was significantly higher in VSCC compared to premalignant lesions, while FasL expression was weak in all categories. However, we suggested the possibility of VSCC cells employing other mechanisms of immunoevasion, such as inhibition of the Fas/FasL apoptotic pathway downstream [41]. Also, Fas receptor has been associated with cell proliferation, mobility, metastasis, and apoptosis resistance, further complicating attempts to elucidate its role in malignant conditions [43]. Future research may explore Fas receptor as a potential target in vulvar malignancies once its biological function is further clarified.
Discussion
Corticotropin-releasing hormone and related peptides play a major role in regulating the stress response system in mammals [44]. Nevertheless, their expression is also found in the peripheral tissues, acting as autocrine and/or paracrine regulators of various biological functions both in healthy tissues and malignant cells [7, 35, 37, 45]. Their exact role in malignancies remains controversial. Herein we aimed to explore the expression and potential implications of CRH family peptides in gynaecological malignancies. Analysing 23 studies in total provided valuable insights into their involvement in various gynaecological cancers. However, it is important to critically evaluate findings and consider their limitations. First of all, 2 types of CRH receptors, CRHR1 and CRHR2, have been characterized, with CRHR1 having a 10-fold higher affinity for CRH [46, 47]. Thus, variations in peptide and receptor concentrations, as well as potential differences in behaviour between in vitro and in vivo studies, can lead to contradictory findings. Therefore, it is crucial to study CRH family peptides and their effects on different malignancies separately and interpret the results cautiously.
Taken together, breast cancer has been extensively studied regarding CRH and related peptides, but the results are inconclusive. Malignant cells in various breast cancer types generally express CRH, urocortin, and their receptors. Some studies suggest that CRH promotes proliferation and inhibits apoptosis, while enhancing cell motility and invasiveness through CRHR1 activation, ultimately favouring metastasis. However, no associations with patient histopathological indices or disease stage have been reported. Other studies indicate that both CRH and urocortin mediate apoptosis and inhibit metastasis through disruption of TGFβ1 signalling. The inconsistent results of studies may stem from variations in experimental conditions, breast cancer subtypes, and tumour microenvironment. The role of CRH family peptides in breast cancer needs further clarification; however, existing data strongly suggest their involvement in tumour growth, progression, and metastasis.
In endometrial malignancy, the expression of CRH, UCNs, and their receptors is well established, with preliminary evidence suggesting their involvement in tumourigenesis. Corticotropin-releasing hormone expression and CRHR1 activation may lead to more aggressive carcinomas with higher migration and metastatic potential, whereas UCN seems to exert a rather protective role via CRHR2 activation, inhibiting cell proliferation and migration. The expression of CRH/UCN in endometrial malignancies appears to be regulated by oestrogens through ERA receptor activation and vice-versa. However, caution is necessary when interpreting these results due to variations in study quality, methodologies, small sample sizes, and the lack of control groups and detailed clinicopathological information.
Limited studies exist regarding human ovarian cancer; nevertheless, consistent upregulation of CRH family peptides, including CRH, CRHR1, CRHR2, and UCN, has been observed. This aligns with previous findings suggesting their involvement in ovarian cancer aggressiveness. Elevated CRH expression is associated with tumour cell proliferation, migration, and invasion, indicating a potential therapeutic target. Furthermore, CRH-induced FasL overexpression may promote immune privilege of tumoural cells, increasing their survivability and progression potential. Specifically, in gynaecological malignancies, expression of both Fas and FasL seems to be associated with tumour growth promotion and metastasis, and a worse overall prognosis [43, 48–51]. Nonetheless, most studies are cross-sectional or retrospective, limiting the ability to establish causal relationships. Prospective studies with larger sample sizes and longitudinal follow-up are needed to determine the prognostic and predictive value of CRH family peptides in ovarian cancer.
Similarly, human cervical cancer cells express CRH and CRHR2, but studies on urocortin expression are lacking. CRH expressed by malignant cells may counterattack the host’s innate immune system by inducing FasL expression or decoy receptor overexpression for NK cells, rendering immunosurveillance ineffective. However, the included studies share similar weaknesses, with small sample sizes and a lack of functional analyses, limiting the understanding of the role of CRH family peptides in cervical cancer immune escape and growth.
Corticotropin-releasing hormone gradually overexpresses from lichen and VIN to vulvar carcinomas, in line with previous results in skin cancer, compared to cutaneous premalignant lesions [52]. This overexpression potentially induces angiogenesis and tumour cell migration [19, 53]. Fas ligand expression also increases progressively, resembling findings in cervical malignancy and CIN [54]. Thus, it could be hypothesized that local inflammation associated with cancer progression may upregulate CRH expression, favouring immune escape [36]. More studies are needed to comprehensively understand the involvement of CRH family peptides in vulvar, fallopian tube, and vaginal malignancies.
Corticotropin-releasing hormone family peptides – CRH, CRHR1, CRHR2, and UCN – are upregulated in gynaecological malignancies, indicating their potential as therapeutic targets and biomarkers. Targeting these peptides and receptors, alone or in combination with other treatment modalities, may disrupt the emerging pro-tumourigenic effects of CRH signalling. Strategies like RNA interference or antibody-based therapies could suppress tumour growth. Nevertheless, systemic effects and physiological processes regulated by CRH family peptides should be considered. Besides, assessing the expression profiles of CRH family peptides in tumour tissues or biological fluids can aid in patient stratification, treatment selection, and monitoring treatment response. Possible correlations with clinicopathological parameters can also provide insights into disease progression and prognosis in gynaecological malignancies.
Despite the potential, various challenges and limitations must be addressed. Achieving selectivity and specificity of agents targeting CRH family peptides, understanding possible resistance mechanisms, and conducting large-scale prospective studies to evaluate efficacy and safety, along with optimal dosage, treatment duration, and combination strategies are crucial for successful and safe implementation of such targeted therapies in clinical practice.
Lastly, future research should focus on understanding the molecular mechanisms and signalling pathways involved in gynaecological malignancies. Identification of reliable biomarkers for patient selection, treatment monitoring, and prognosis prediction is essential. Preclinical and clinical investigations should explore combination therapies, such as immunotherapy or targeted agents, to uncover potential synergistic effects. By addressing these areas, we can advance our understanding of gynaecological cancer biology and develop improved therapeutic strategies for better patient outcomes.
Conclusions
Studying the current literature, it becomes apparent that CRH family peptides and their receptors are not only expressed, but also play a significant role, in female malignancies. Whether this role can be utilized for the patient’s benefit is a subject of future clinical studies with CRH receptor agonists/antagonists. To date, mostly based on in vitro preliminary studies, it seems that CRH/CRHR1 activation in female malignancies favours cancer cell motility and migration potential and attenuation of the host’s innate immune response to cancer cells. Urocortin and CRHR2 activation, on the other hand, seems to play a protective role, especially in endometrial cancer, through cell proliferation and migration inhibition. An exception seems to be breast cancer, in which results are conflicting. Nevertheless, the promising results from preliminary studies reviewed herein should prompt for further research on CRH family peptides in gynaecological tumourigenesis. Clearly, more clinically oriented studies with larger samples are needed to investigate whether an iatrogenic intervention with specific CRH receptor agonists or antagonists would benefit female cancer patients.








