

Introduction
Atherosclerosis (AS) is a common vascular disease characterized by lipid and fibrous elements amassing in the wall of the artery [1]. The pathogenesis of AS is connected with elevated oxidative stress, expression of chemokines/cytokines, and activation of pro-inflammatory signaling pathways [2]. Also, a crucial factor in the progression of AS is oxidized low-density lipoprotein (ox-LDL) [3]. Ox-LDL participated in AS mainly by inducing apoptosis and oxidative stress of endothelial cells and destroying the antioxidant and secretory activity of the vascular endothelium [4]. Consequently, it is indispensable to probe into the potential molecular mechanisms of AS to provide possible strategies for preventing or treating AS.
Circular RNAs (circRNAs) are a type of covalent cyclic endogenous RNAs that are formed by the covalent attachment to the 5' splice acceptor in the reverse order by a 3' splice donor of pre-mRNA [5]. A few possible features have been reported: binging protein, miRNA sponge, and translation regulators [6]. Moreover, circRNA was revealed to be connected with many diseases, including AS [7]. For example, circular RNA hsa_circ_0044073 expedited cell proliferation and invasion in AS [8]. Circular RNA hsa_circ_0003204 (hsa_circ_0003204) originated from chromosomal region 17q25.3 and was an exon transcript of ubiquitin-specific peptidase 36 (USP36). Hsa_circ_0003204 was found to be enhanced in human umbilical vein endothelial cells treated with ox-LDL [9]. However, the role of hsa_circ_0003204 and its molecular mechanism in AS are unclear.
MicroRNAs (miRNAs) are an excellent class of non-coding RNAs (approximately 22 nucleotides) that exert their functions primarily via translational inhibition or mRNA degradation for the post-transcriptional regulation of gene expression [10]. MicroRNA-330-5p (miR-330-5p) has been revealed to be associated with AS progression [11], carotid plaque’s stability [12] and tumor occurrence [13, 14]. Nevertheless, whether miR-330-5p is regulated by hsa_circ_0003204 in the progression of AS has not been reported.
The nucleotide-binding oligomerization domain 2 (Nod2), which is rich in leucine repeats, is a member of the nucleotide-binding and oligomerization domain (NOD)-like receptor (NLR) family [15]. It is a major recognition receptor that integrates inflammation and endoplasmic reticulum stress and plays crucial roles in tissue homeostasis and innate antibacterial immunity [16]. It was claimed that Nod2 plays vital roles in the inflammatory process of AS [17]. Also, Nod2 was revealed to be connected with endothelial cell reactive oxygen species (ROS) generation [18] and injury [19, 20]. However, the molecular mechanisms associated with Nod2 in AS have rarely been reported.
Hence, we investigated the effect of ox-LDL on the viability of human primary aortic endothelial cells (HAECs) and constructed an AS cell model. Moreover, the expression pattern and role of hsa_circ_0003204 in the AS cell model were probed. Furthermore, the mechanism of the hsa_circ_0003204/miR-330-5p/Nod2 axis in HAEC cells was investigated, providing possible tactics for the prevention or treatment of AS.
Material and methods
Cell culture
Human primary aortic endothelial cells (HAECs) were procured from American Tissue Culture Collection (Manassas, VA, USA). An incubator in a humidifier with 5% CO2 at 37°C was utilized to cultivate HAECs. Roswell Park Memorial Institute (RPMI) 1640 medium (HyClone, Logan, UT, USA) supplemented with fetal bovine serum (10%, FBS, Atlanta Biologicals, Atlanta, GA) was applied to culture HAECs. Synchronously, streptomycin (1%, Baomanbio, Shanghai, China) and penicillin (1%, Baomanbio) were replenished to the RPMI 1640 medium.
Cell treatment
HAECs were cultured in RPMI 1640 medium with different concentrations of ox-LDL (10, 20, 40 µg/ml) for 24 h in order to evaluate the effect of ox-LDL on the viability of HAECs. Also, HAECs were cultured in RPMI 1640 medium with ox-LDL (20 µg/ml) for 24 h, 48 h, and 72 h to assess the effect of ox-LDL on HAEC viability. HAECs with ox-LDL (20 µg/ml) treatment for 48 h were used as an AS cell model for further studies.
Cell transfection
Small interfering RNA targeting hsa_circ_0003204 (si-hsa_circ_0003204-1 and si-hsa_circ_0003204-2) and negative control (si-NC) were obtained from Genepharma (Shanghai, China). The miRNA mimic or inhibitor targeting miR-330-5p (miR-330-5p or in-miR-330-5p) and their corresponding negative control (miR-NC or in-miR-NC) were obtained from Ambion Inc. (Austin, TX, USA). The pcDNA3.1 vector (Invitrogen, Carlsbad, CA, USA) was applied to establish the pcDNA-Nod2 overexpression vector (Nod2). Lipofectamine 3000 reagent (Invitrogen) was utilized for the transfection of ox-LDL-induced HAECs.
Cell viability
The viability of HAECs with or without ox-LDL treatment was determined counting kit-8 (CCK8) assay. In short, HAECs (2 × 104) were maintained in medium with different concentrations of ox-LDL (10, 20, 40 µg/ml) for 24 h or were maintained in medium with ox-LDL (20 µg/ml) for 24 h, 48 h, and 72 h. Then, CCK8 (10 µl, Dojindo Laboratories, Tokyo, Japan) was replenished to each well for 2 h. Following this, the color reaction at 450 nm was determined through the Microplate Absorbance Reader (Thermo Fisher Scientific, Waltham, MA, USA).
Reactive oxygen species, malondialdehyde and superoxide dismutase measurement
The Cellular Reactive Oxygen Species Detection Assay Kit (Abcam, Cambridge, MA, USA) and Infinite 200 PRO Microplate Reader (Sunrise, Tecan, Switzerland) were used for detection of the level of ROS in the supernatant of ox-LDL-induced HAECs with or without transfection. The Lipid Peroxidation Assay Kit (Abcam) was utilized for evaluation of the level of malondialdehyde (MDA) in the supernatant of ox-LDL-induced HAECs with or without transfection, and the Infinite 200 PRO Microplate Reader was used to determine the color reaction at 532 nm. The Superoxide Dismutase Activity Assay Kit (Abcam) was applied to analyze the level of superoxide dismutase (SOD) in the supernatant of ox-LDL-induced HAECs with or without transfection, and the color reaction at 450 nm was determined by the Infinite 200 PRO Microplate Reader.
Flow cytometry assay
The apoptosis rate of ox-LDL-induced HAECs was assessed through the Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection kit (BD Biosciences, San Jose, CA, USA). Briefly, ox-LDL-induced HAECs with or without transfection were cultured in the medium for 48 h. After washing, the cells were re-suspended in binding buffer (200 µl) at a concentration of 2 × 105 cells/ml. Following this, the cells were stained with Annexin V-FITC (5 µl) and PI (100 µl) for 30 min in the dark. Eventually, the apoptosis rate of ox-LDL-induced HAECs with or without transfection was assessed through the FACScan flow cytometry (BD Biosciences).
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA of HAECs and ox-LDL-induced HAECs with or without transfection was extracted through TRIzol reagent (Thermo Fisher Scientific). Moloney Murine Leukemia Virus (M-MLV) First Strand Kit (Life Technologies, Grand Island, NY, USA) or One Step PrimeScript miRNA cDNA Synthesis Kit (Takara, Dalian, China) was utilized for generation of the first-strand complementary DNA of hsa_circ_0003204, Nod2, and miR-330-5p. The SYBR Fast qPCR Mix (Thermo Fisher Scientific) was used to analyze the levels of hsa_circ_0003204, miR-330-5p and Nod2. U6 small nuclear RNA (snRNA) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was acted as an internal control for miR-330-5p, hsa_circ_0003204, and Nod2. The levels of hsa_circ_0003204, miR-330-5p, and Nod2 were calculated by the 2-ΔΔCt method. Primers for hsa_circ_0003204 were purchased from Sangon Biotech (Shanghai, China). The primers used were as follows: miR-330-5p: 5'-TCTCTGGGCCTGTGTCTTAGGC-3' (F) and 5'-TTAATGGGGTGATTGGTGGT-3' (R); Nod2: 5'-CCAGCGTCTTTGGCCATTCAACAT-3' (F) and 5'-TTGAGCTCATCCAGTGCTTGGAGT-3' (R); GAPDH: 5'-GACTCCACTCACGGCAAATTCA-3' (F) and 5'-TCGCTCCTGGAAGATGGTGAT-3' (R), Lectin-like ox-LDL receptor-1 (LOX-1): 5'-TTACTCTCCATGGTGGTGCC-3' (F) and 5'- AGCTTCTTCTDCTTGTTGCC-3' (R) as well as U6 snRNA: 5'-GCTCGCTTCGGCAGCACA-3' (F) and 5'-GAGGTATTCGCACCAGAGGA-3'(R).
Dual-luciferase reporter assay
The CircInteractome database or DIANA microT-CDS database was employed for prediction of the binding sites between hsa_circ_0003204 or Nod2 and miR-330-5p. The sequences of hsa_circ_0003204 and Nod2 3' untranslated regions (UTR) (with predicted miR-330-5p binding sites) or their mutants were amplified and embedded into the pGL3 vector (Promega, Fitchburg, WI, USA) for construction of the luciferase reporter vector of wild-type hsa_circ_0003204 (hsa_circ_0003204 WT) and wild-type Nod2 3'UTR (Nod2 3'UTR WT) as well as mutant hsa_circ_0003204 (hsa_circ_0003204 MUT) and mutant Nod2 3'UTR (Nod2 3'UTR MUT). Next, the luciferase reporter vectors were transfected into HAECs with miR-330-5p, miR-NC, in-miR-330-5p, or in-miR-NC using Lipofectamine 3000 transfection reagent (Invitrogen). In the end, the dual-luciferase reporter assay kit (Promega) was used to evaluate the luciferase activities of the luciferase reporter vectors in HAECs.
Western blot analysis
Total protein of ox-LDL-induced HAECs with transfection was extracted with radio-immunoprecipitation assay (RIPA) lysis buffer (Thermo Fisher Scientific). Next, total protein was segregated with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (8-12%, SDS-PAGE). Following that, the wet electrophoretic transfer method was executed for transference of the segregated protein to polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA, USA). Afterward, tris buffered saline tween (TBST) buffer with 5% skim milk was employed to block the PVDF membranes. After that, the PVDF membranes were incubated with primary antibodies overnight at 4°C. The antibodies were as follows: anti-Nod2 (ab31488, 1 : 1000, Abcam), anti-LOX-1 (ab60178, 1 : 1000, Abcam), and anti-β-actin (ab11003, 1 : 1000, Abcam). Then, the membranes were incubated with goat anti-mouse (ab205719, 1 : 2000, Abcam) or rabbit (ab205718, 1 : 2000, Abcam) IgG. β-actin was regarded as a loading control. Finally, the bands were visualized with the Immobilon Western Chemiluminescent HRP Substrate (Millipore).
Statistical analysis
The data in the study were derived from at least 3 independent experiments. GraphPad Prism 6.0 (GraphPad, San Diego, CA, USA) and SPSS 20.0 software (IBM, Armonk, NY, USA) were used for statistical analysis. The differences between two groups were analyzed through Student’s t test. Data are shown as mean ± standard deviation. Differences with p < 0.05 were statistically significant.
Results
Hsa_circ_0003204 was elevated in ox-LDL-induced HAECs
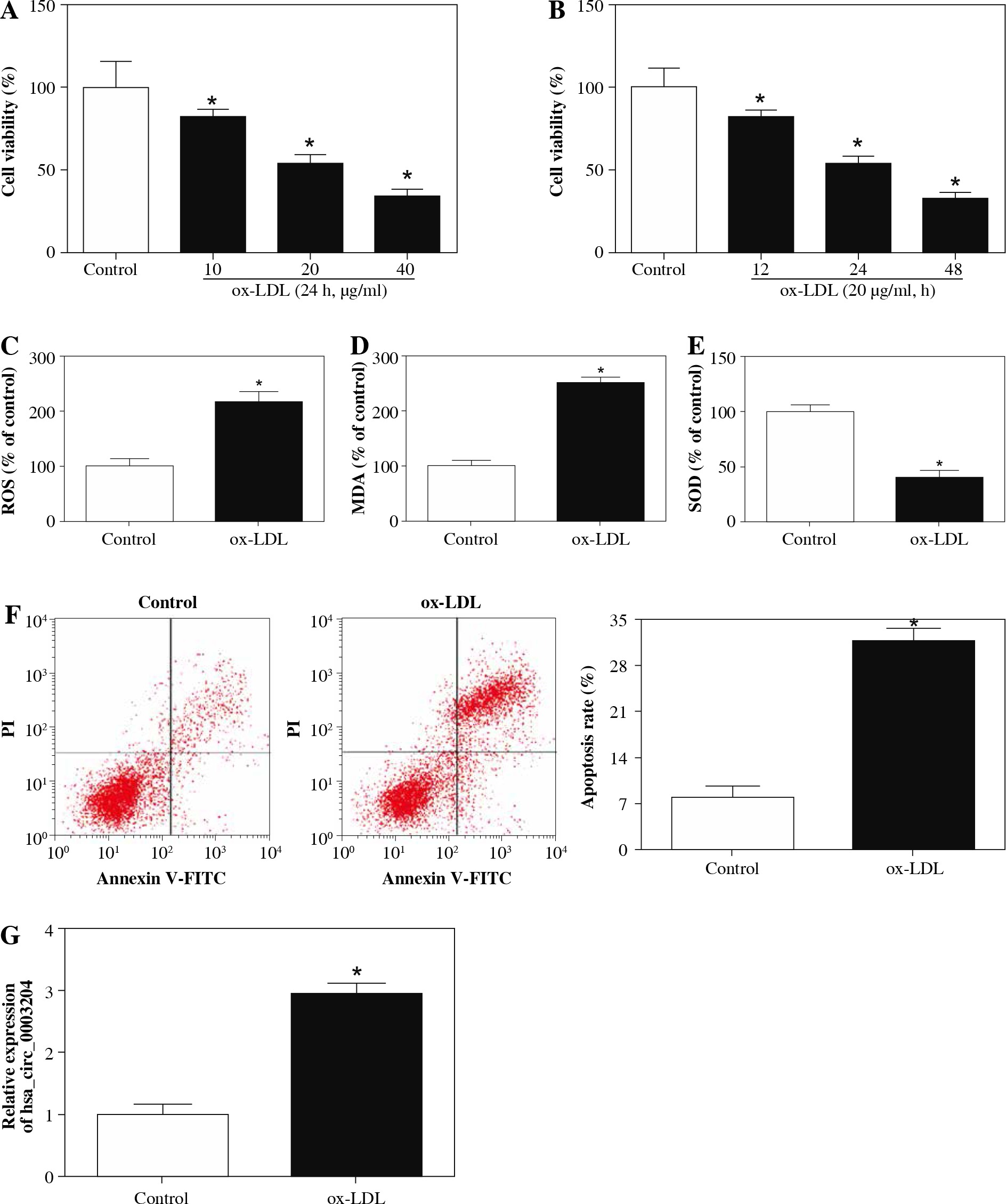
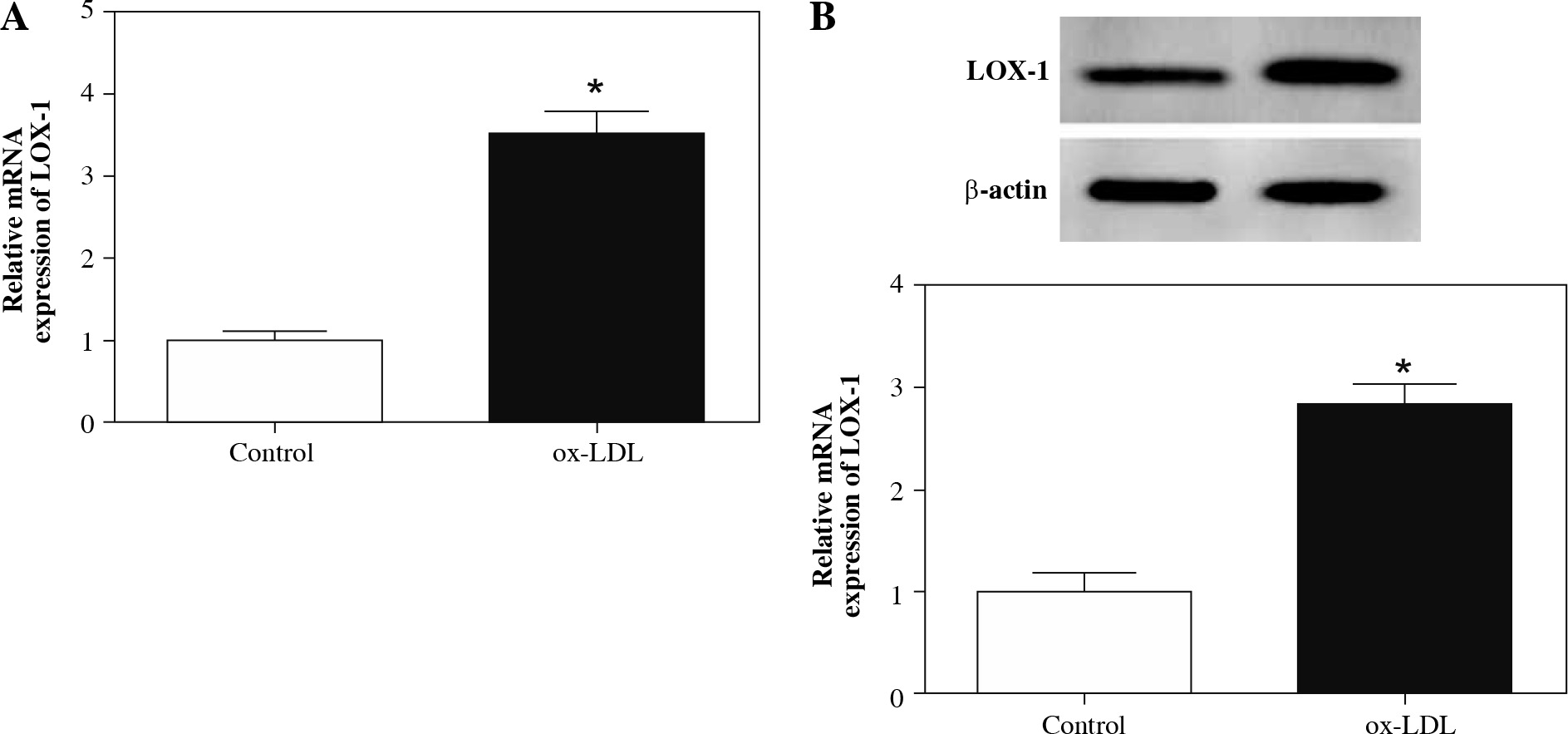
In the first place, RPMI 1640 medium with different concentrations of ox-LDL (10, 20, 40 µg/ml) was utilized to test the effect of ox-LDL on the viability of HAECs. Results of CCK-8 showed that the viability of HAECs induced by ox-LDL was evidently repressed in a concentration-dependent manner compared with the control group (Fig. 1A). Moreover, the viability of HAECs treated with ox-LDL (20 µg/ml) for 12 h, 24 h, 48 h was determined with CCK-8 assay. The results showed that the viability of HAECs treated with ox-LDL was dramatically constrained in a time-dependent manner (Fig. 1B). Also, HAECs were induced with ox-LDL (20 µg/ml) for 48 h for subsequent studies. Next, we assessed the oxidative stress and apoptosis of ox-LDL-induced HAECs. The results showed that the levels of ROS and MDA in the supernatant of ox-LDL-induced HAECs were elevated, but the level of SOD was impeded (Fig. 1C-E). Flow cytometry assay revealed that the apoptosis rate of ox-LDL-induced HAECs was remarkably higher than that in the control group (Fig. 1F). LOX-1 is a receptor for ox-LDL, which regulates the biological function of ox-LDL. We observed that the levels of LOX-1 mRNA and protein were elevated in ox-LDL-induced HAECs (Fig. 2A and B). Therefore, the induction of HAECs with ox-LDL (20 µg/ml) for 48 h was used as an AS cell model for further study. Subsequently, the expression of hsa_circ_0003204 in ox-LDL-induced HAECs was evaluated by qRT-PCR. As exhibited in Figure 1G, the level of hsa_circ_0003204 was apparently higher in ox-LDL-induced HAECs compared to the control group. In sum, high hsa_circ_0003204 expression might be connected with the process of AS.
Fig. 1
Effect of ox-LDL on expression of hsa_circ_ 0003204 in HAECs. A) CCK-8 assay was conducted to determine the viability of HAECs treated with ox-LDL (10, 20, 40 μg/ml) for 24 h. B) CCK-8 assay was carried out for assessment of the viability of HAECs treated with ox- LDL (20 μg/ml) for 12 h, 24 h, 48 h. C-E) Levels of ROS, MDA and SOD in the supernatant of ox-LDL-induced HAECs were analyzed with the microplate reader. F) Flow cytometry assay was employed to detect apoptosis of ox- LDL-induced HAECs. G) qRT-PCR was applied to assess the expression of hsa_circ_0003204 in ox-LDL-induced HAECs. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
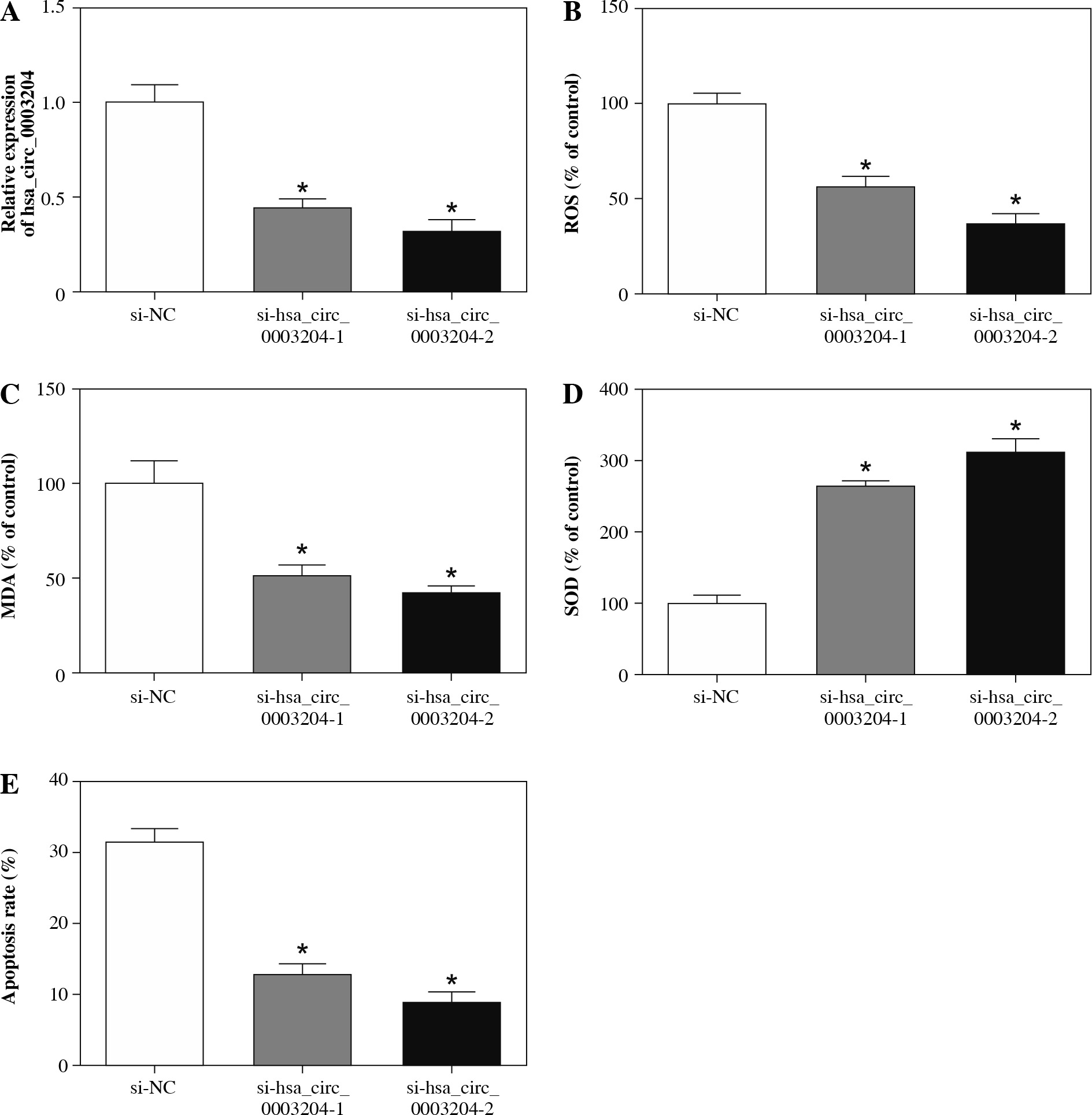
Hsa_circ_0003204 depletion constrained the oxidative stress and apoptosis of ox-LDL-induced HAECs
Taking into account the above results, we further probed into the role of hsa_circ_0003204 in the process of AS by loss-of-function experiments. In the first place, the si-NC, si-hsa_circ_0003204-1, or si-hsa_circ_0003204-2 was transfected into ox-LDL-induced HAECs to silence the expression of hsa_circ_0003204. Results of qRT-PCR indicated that hsa_circ_0003204 was obviously reduced in ox-LDL-induced HAECs transfected with si-hsa_circ_0003204-1 and si-hsa_circ_0003204-2 in contrast to the si-NC group (Fig. 3A). Afterward, the effects of hsa_circ_0003204 inhibition on the oxidative stress and apoptosis of ox-LDL-induced HAECs were explored. The results demonstrated that ROS and MDA were conspicuously suppressed, while SOD was distinctly accelerated in ox-LDL-induced HAECs transfected with si-hsa_circ_0003204-1 and si-hsa_circ_0003204-2 compared with the control group (Fig. 3B-D). Also, flow cytometry revealed that si-hsa_circ_0003204-1 and si-hsa_circ_0003204-2 could drastically impede the apoptosis of ox-LDL-induced HAECs vs. the control si-NC (Fig. 3E). Taken together, these results suggested that inhibition of hsa_circ_0003204 could attenuate ox-LDL induced injury of HAECs by repressing oxidative stress and apoptosis.
Fig. 3
Effects of hsa_circ_0003204 on oxidative stress and apoptosis of ox-LDL-induced HAECs. A-E) HAECs treated with ox-LDL were transfected with si-NC, si-hsa_ circ_0003204-1 or si-hsa_circ_0003204-2. A) Expression of hsa_circ_0003204 in HAECs treated with ox-LDL was analyzed by qRT-PCR. B-D) Levels of ROS, MDA and SOD in supernatant of HAECs treated with ox-LDL were evaluated through the microplate reader. E) Apoptosis rate of HAECs treated with ox-LDL was analyzed via flow cytometry assay. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
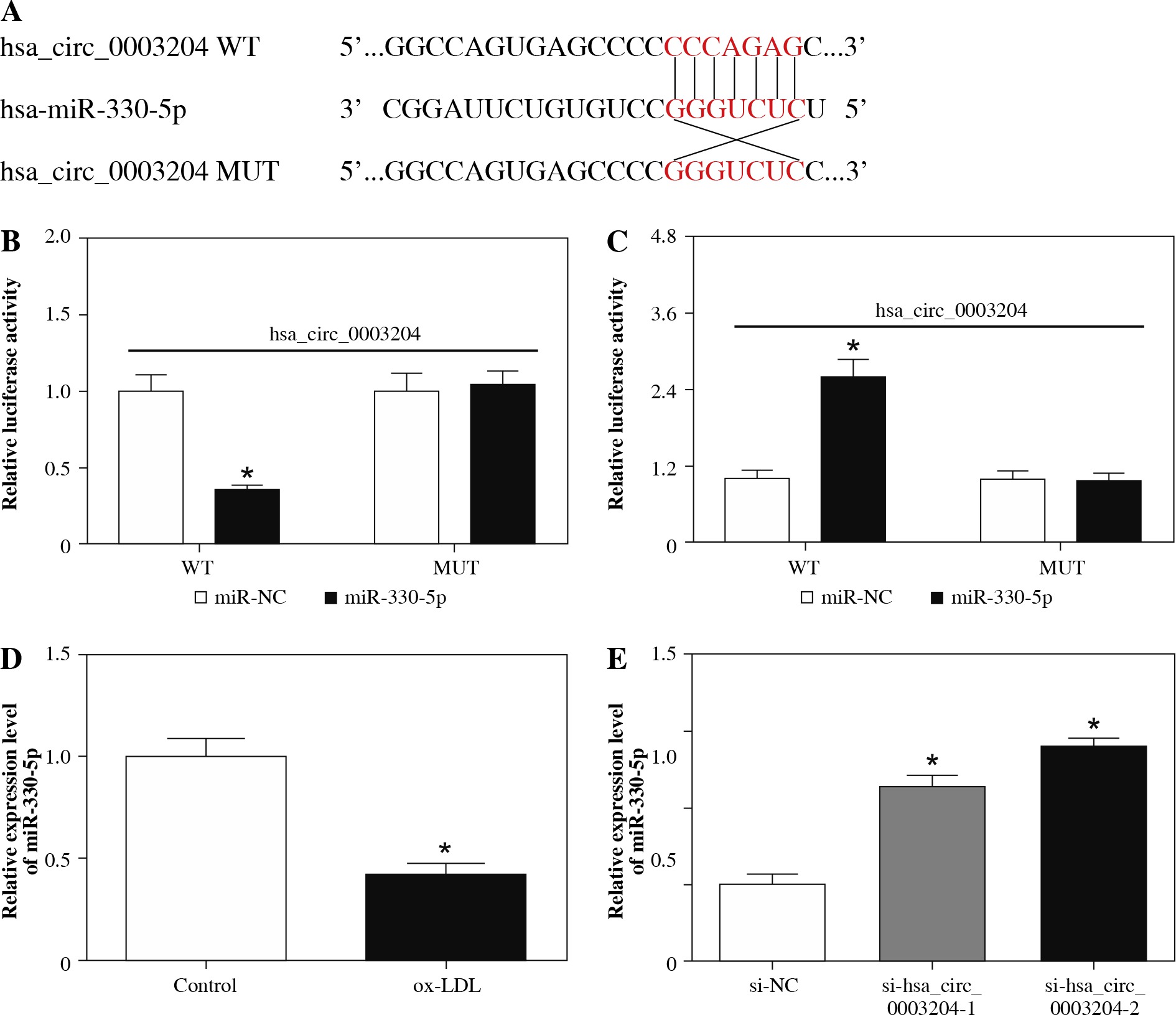
Hsa_circ_0003204 acted as a sponge of miR-330-5p
To further understand how hsa_circ_0003204 played its function in AS, the CircInteractome database was used for the prediction of the target of hsa_circ_0003204. We discovered that miR-330-5p possessed potential binding sites for hsa_circ_0003204 (Fig. 4A). Afterward, we constructed luciferase reporter vectors with hsa_circ_0003204 WT and hsa_circ_0003204 MUT to execute the dual-luciferase reporter assay for the verification of binding sites between hsa_circ_0003204 and miR-330-5p. The results showed that the luciferase activity of the luciferase reporter vectors containing hsa_circ_0003204 WT was strikingly lower in HAECs transfected with miR-330-5p than the miR-NC group (Fig. 4B). Moreover, silenced miR-330-5p expression enhanced the luciferase activity of the luciferase reporter vectors containing hsa_circ_0003204 WT in HAECs compared to the vector control group (Fig. 4C). Additionally, qRT-PCR was conducted to detect miR-330-5p expression in ox-LDL-induced HAECs. The results showed that miR-330-5p was dramatically downregulated in ox-LDL-induced HAECs (Fig. 4D). Also, a conspicuous augmentation of miR-330-5p was observed in ox-LDL-induced HAECs transfected with si-hsa_circ_0003204-1 and si-hsa_circ_0003204-2 (Fig. 4E). In short, these findings demonstrated that hsa_circ_0003204 served as a sponge of miR-330-5p in ox-LDL-induced HAECs.
Fig. 4
MiR-330-5p was a target of hsa_circ_0003204 in HAECs. A) Binding sites between miR-330-5p and hsa_ circ_0003204 were predicted by the CircInteractome database. B, C) Activities of luciferase reporter vectors (hsa_ circ_0003204 WT and hsa_circ_0003204 MUT) in HAECs transfected with miR-NC (B), miR-330-5p (B), in-miR-NC, (C) or in-miR-330-5p (C) were evaluated with dual-luciferase reporter assay. D) Expression of miR-330-5p in ox-LDLinduced HAECs was detected by qRT-PCR. E) Level of miR-330-5p in ox-LDL-induced HAECs transfected with si- NC, si-hsa_circ_0003204-1, or si-hsa_circ_0003204-2 was analyzed via qRT-PCR. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
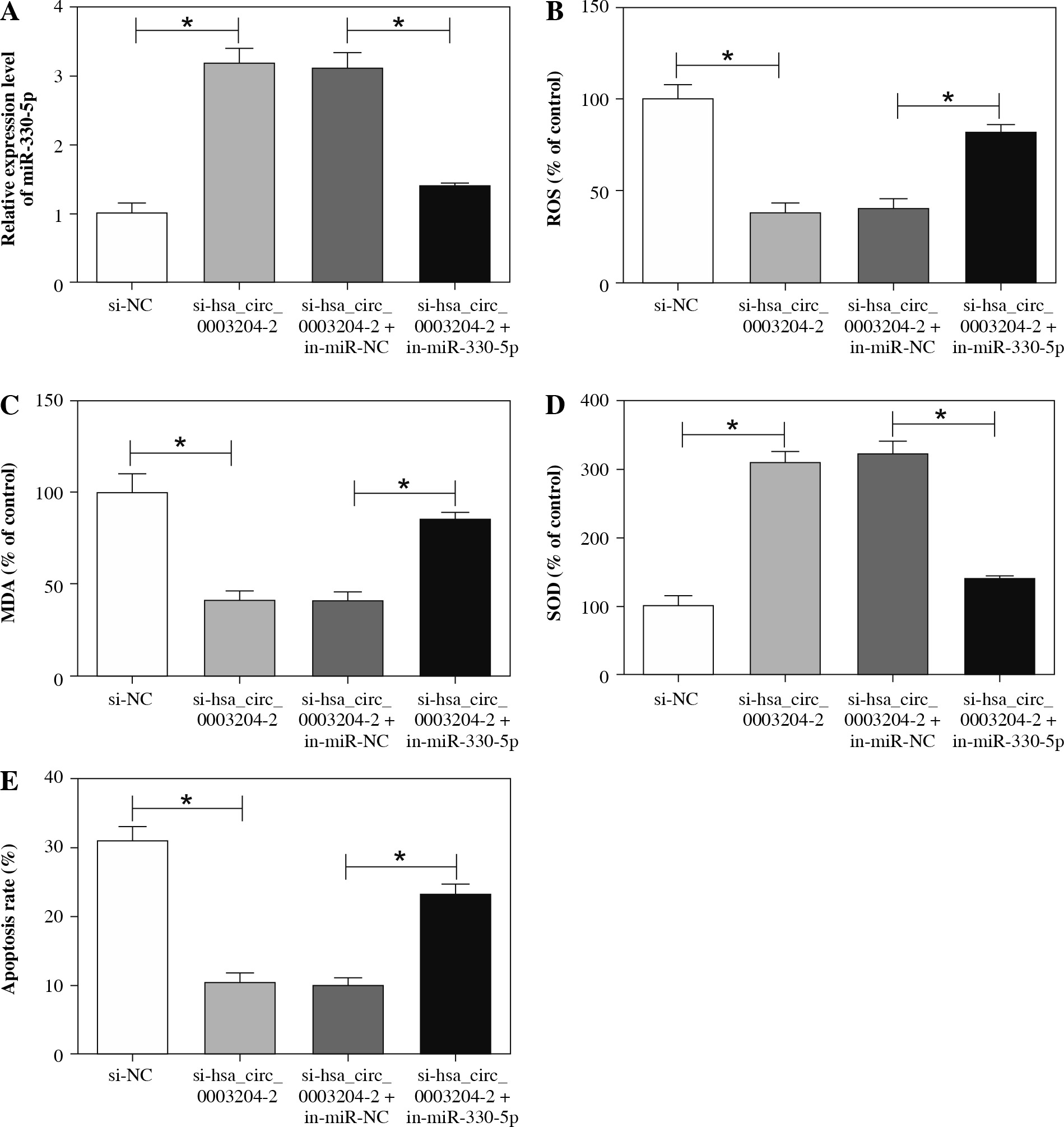
MiR-330-5p silencing reversed hsa_circ_0003204 knockdown-mediated effects on cell oxidative stress and apoptosis in ox-LDL-induced HAECs
In view of the above results, we further explored whether hsa_circ_0003204 mediated the oxidative stress and apoptosis of ox-LDL-induced HAECs via miR-330-5p. At the outset, qRT-PCR was carried out to detect the expression of miR-330-5p in ox-LDL-induced HAECs transfected with si-NC, si-hsa_circ_0003204-2, si-hsa_circ_0003204-2 + in-miR-NC, or si-hsa_circ_0003204-2 + in-miR-330-5p. The results indicated that the enhancement of miR-330-5p in ox-LDL-induced HAECs caused by hsa_circ_0003204 knockdown was abolished by the inhibition of miR-330-5p (Fig. 5A). Following this, the effects of miR-330-5p suppression on the oxidative stress and apoptosis of ox-LDL-induced HAECs transfected with si-hsa_circ_0003204-2 were further evaluated. The results showed that both the reduction of ROS and MDA and the augmentation of SOD in ox-LDL-induced HAECs caused by hsa_circ_0003204 downregulation were overturned by the inhibition of miR-330-5p (Fig. 5B-D). Flow cytometry indicated that decreased miR-330-5p expression could restore hsa_circ_0003204 repression-mediated effects on cell oxidative stress and apoptosis in ox-LDL-induced HAECs (Fig. 5E). Therefore, these results demonstrated that hsa_circ_0003204 regulated cell oxidative stress and apoptosis in ox-LDL-induced HAECs through miR-330-5p.
Fig. 5
Hsa_circ_0003204 worked via regulating miR-330-5p expression in ox-LDL-induced HAECs. A-E) Ox-LDL-induced HAECs were transfected with si-NC, si-hsa_circ_0003204-2, si-hsa_circ_0003204-2 + in-miR-NC or si-hsa_circ_0003204-2 + in-miR-330-5p. A) Expression of miR-330-5p was analyzed through qRT-PCR. B-D) The microplate reader was used to analyze the levels of ROS, MDA, and SOD in supernatant of ox-LDL-induced HAECs. E) Flow cytometry assay was applied to detect the apoptosis rate of ox-LDL-induced HAECs. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
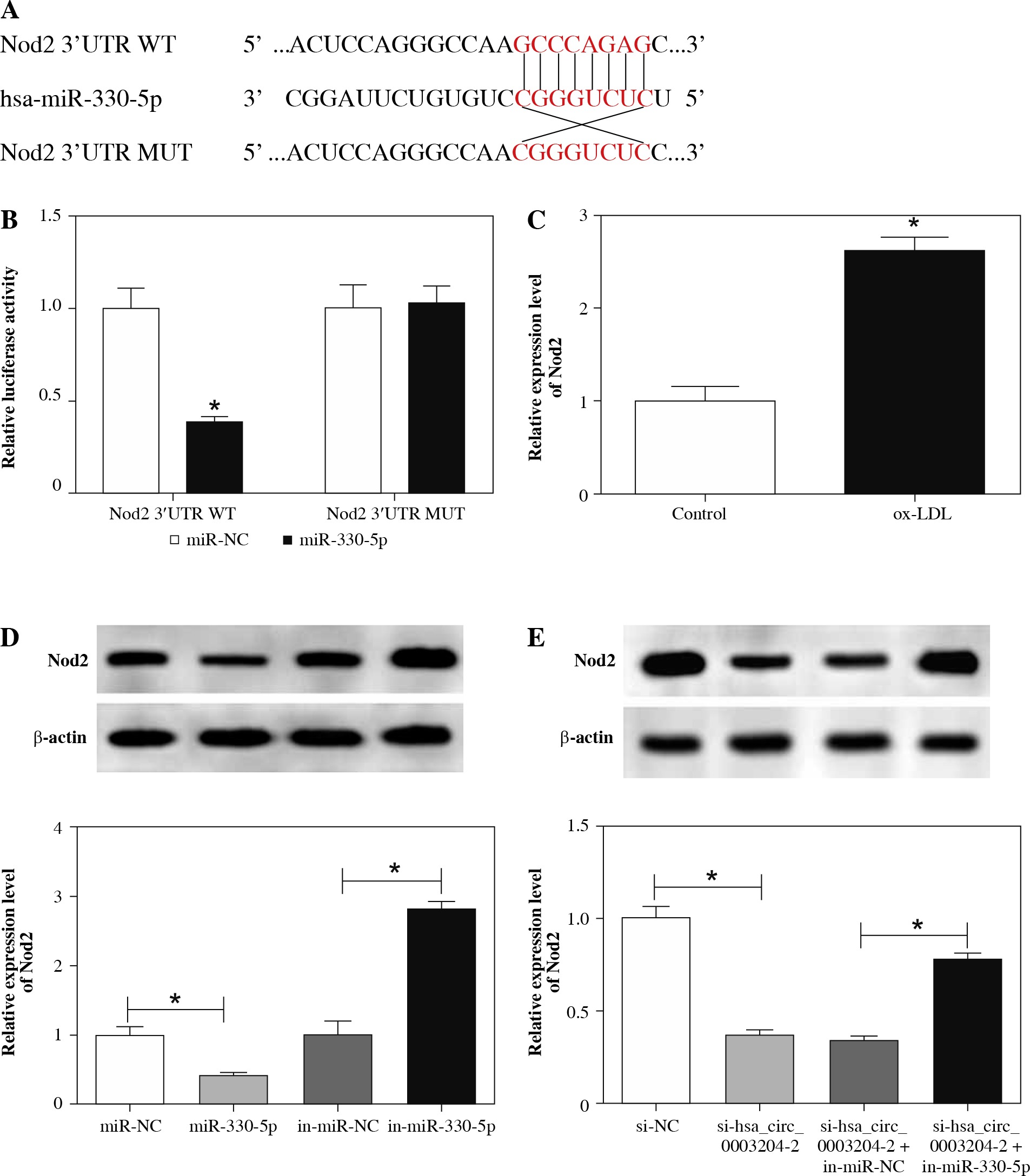
Nod2 was verified to act as a target of miR-330-5p in HAECs
To figure out how miR-330-5p exerted its role in AS, the DIANA microT-CDS database was employed to predict the target of miR-330-5p in HAECs. As presented in Figure 6A, Nod2 was predicted to have possible binding sites for miR-330-5p. Then, the luciferase reporter vectors with Nod2 3'UTR WT and Nod2 3'UTR MUT were established. The results of dual-luciferase reporter assay revealed that miR-330-5p specially reduced the activity of the luciferase reporter vectors containing Nod2 3'UTR WT in HAECs, while the activity of the luciferase reporter vectors containing Nod2 3'UTR MUT was not apparently different (Fig. 6B). Moreover, results of qRT-PCR showed that the mRNA level of Nod2 was remarkably boosted in ox-LDL-induced HAECs (Fig. 6C). Also, the protein level of Nod2 in ox-LDL-induced HAECs transfected with miR-NC, miR-330-5p, in-miR-NC, or in-miR-330-5p was analyzed with western blot analysis. As expected, miR-330-5p overexpression dramatically decreased the protein level of Nod2 in ox-LDL-induced HAECs, whereas downregulation of miR-330-5p drastically elevated the Nod2 protein level, indicating that Nod2 was negatively regulated by miR-330-5p (Fig. 6D). Furthermore, we discovered that Nod2 protein level was evidently downregulated in ox-LDL-induced HAECs transfected with si-hsa_circ_0003204-2, while this reduction was recovered by the introduction of in-miR-330-5p, indicating that Nod2 was regulated by hsa_circ_0003204 and miR-330-5p (Fig. 6E). Collectively, these results suggested that Nod2 was regulated by hsa_circ_0003204 via miR-330-5p in HAECs.
Fig. 6
MiR-330-5p targeted Nod2 in HAECs. A) Binding sites of miR-330-5p in Nod2 were predicted by the DIANA microT-CDS database. B) Luciferase activities of Nod2 3'UTR WT and Nod2 3'UTR MUT in HAECs transfected with miR-330-5p or miR-NC were assessed by dual-luciferase reporter assay. C) mRNA level of Nod2 in ox-LDL-induced HAECs was measured through qRT-PCR. D) Protein level of Nod2 in ox-LDL-induced HAECs transfected with miR-NC, miR-330-5p, in-miR-NC, or in-miR-330-5p was evaluated using western blot analysis. E) Western blot analysis was carried out for evaluation of the protein level of Nod2 in ox-LDL-induced HAECs transfected with si-NC, sihsa_ circ_0003204-2, si-hsa_circ_0003204-2 + in-miR-NC, or si-hsa_circ_0003204-2 + in-miR-330-5p. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
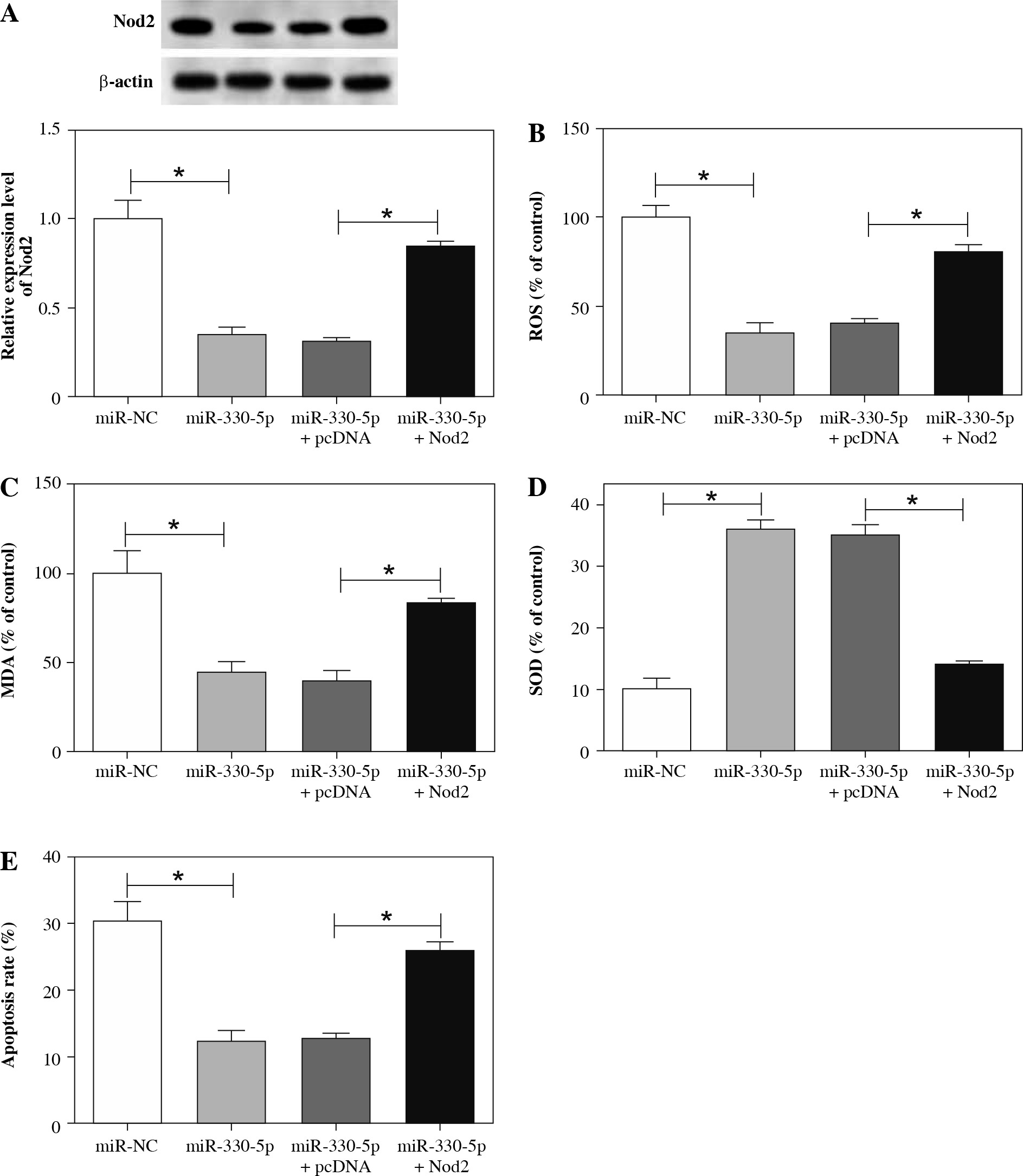
Nod2 augmentation reversed the effects of miR-330-5p on the oxidative stress and apoptosis of ox-LDL-induced HAECs
Knowing that miR-330-5p negatively regulated Nod2 expression, we next investigated whether miR-330-5p mediated the oxidative stress and apoptosis of ox-LDL-induced HAECs via Nod2. Firstly, the protein level of Nod2 in ox-LDL-induced HAECs transfected with miR-NC, miR-330-5p, miR-330-5p + pcDNA, or miR-330-5p + Nod2 was assessed with western blot analysis. The results indicated that the inhibition of Nod2 protein mediated by miR-330-5p upregulation was restored by the overexpression of Nod2 (Fig. 7A). Furthermore, we found that both the constraint of ROS and MDA and the acceleration of SOD in the supernatant of ox-LDL-induced HAECs prompted by miR-330-5p elevation were recovered by Nod2 upregulation (Fig. 7B-D). Moreover, Nod2 enhancement overturned the inhibition of apoptosis of ox-LDL-induced HAECs earned by miR-330-5p elevation (Fig. 7E). Together, these data indicated that miR-330-5p modulated cell oxidative stress and apoptosis in ox-LDL-induced HAECs by Nod2.
Fig. 7
MiR-330-5p regulated Nod2 expression to mediate oxidative stress and apoptosis in ox-LDL-induced HAECs. A-E) Ox-LDL-induced HAECs were transfected with miR-NC, miR-330-5p, miR-330-5p + pcDNA, or miR-330-5p + Nod2. A) Protein level of Nod2 in HAECs with ox-LDL treatment was detected by western blot analysis. B-D) Levels of ROS, MDA, and SOD in the supernatant of HAECs with ox-LDL treatment were analyzed with a microplate reader. E) Apoptosis rate of HAECs with ox-LDL treatment was determined through flow cytometry assay. Data are shown as mean ± standard deviation of 3 independent experiments. *p < 0.05
Discussion
Atherosclerosis is a major health problem for the elderly worldwide, and it can cause cardiovascular diseases such as stroke and coronary heart disease [21]. Studies have shown that circRNA is stable with cell type-specific or tissue-specific expression, and it is a promising disease biomarker [22]. Hsa_circ_0003204 was claimed to be augmented in vascular endothelial cells with ox-LDL treatment, while the role of hsa_circ_0003204 in AS was unclear.
Oxidative stress plays vital roles in AS pathogenesis by participating in the oxidative modification of ox-LDL. Its imbalance reduced the body’s inherent antioxidant defense system or contributed to the production of ROS [23]. Also, an excess of ROS could cause cardiovascular disease [24]. SOD is a vascular wall antioxidant enzyme that converts superoxide to hydrogen peroxide [25]. Elevated serum MDA levels might play a vital role in the development of AS, leading to coronary heart disease [26]. In this study, HAECs were treated with ox-LDL to construct AS models. We discovered that ROS and MDA levels were enhanced in the supernatant of ox-LDL-induced HAECs, while the SOD level was reduced. Moreover, the apoptosis was promoted and the expression of hsa_circ_0003204 was upregulated in HAECs with ox-LDL treatment. Furthermore, silenced hsa_circ_0003204 expression blocked the oxidative stress and apoptosis of ox-LDL-induced HAECs. Nevertheless, little is known about the expression and role of hsa_circ_0003204 in AS and the other diseases.
Studies have revealed that circRNA exerts its function by acting as a sponge of miRNA [27-29]. MiR-330-5p was claimed to be involved in the progression of AS. Wei et al. uncovered that miR-330-5p elevation might cause carotid plaque instability in patients with carotid stenosis [12]. Additionally, miR-330-5p was disclosed to be decreased in THP-1 cells with ox-LDL treatment and miR-330-5p enhancement curbed the process of oxidative stress and inflammatory THP-1 cells with ox-LDL treatment [30]. In the present study, hsa_circ_0003204 was confirmed to serve as a sponge of miR-330-5p in HAECs. Furthermore, consistent with the above studies, miR-330-5p was downregulated in ox-LDL-induced HAECs and miR-330-5p repression reversed the suppression of oxidative stress and apoptosis of ox-LDL-induced HAECs caused by hsa_circ_0003204 knockdown.
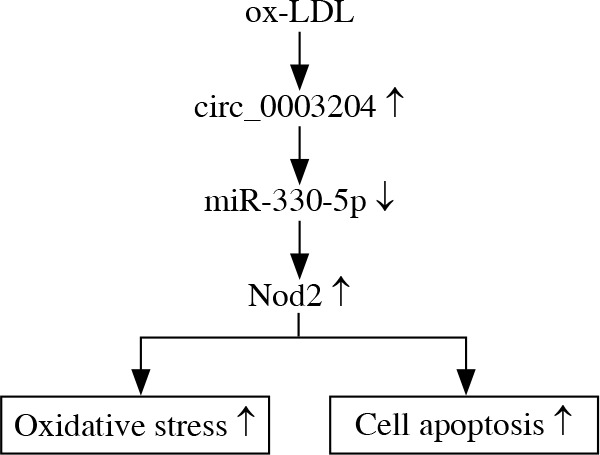
It was reported that Nod2 could increase lipid-rich necrotic core expression and contribute to vascular inflammation in AS [31]. Moreover, AS was associated with Nod2-mediated increase in innate immunity [16]. The report of Xu et al. revealed that Nod2 downregulation could mitigate endothelial cell injury induced by ox-LDL [19]. Also, the reduction of Nod2 caused by long noncoding RNA XIST inhibition could attenuate endothelial cell injury by repressing apoptosis and promoting the viability of ox-LDL-induced umbilical vein endothelial cells [20]. In this study, we discovered that Nod2 was enhanced in ox-LDL-induced HAECs and Nod2 was the target of miR-330-5p. Moreover, the introduction of Nod2 abolished miR-330-5 upregulation-mediated oxidative stress and apoptosis of ox-LDL-induced HAECs. In addition, hsa_circ_0003204 regulated Nod2 protein via miR-330-5p in HAECs. Therefore, we verified that Nod2 could accelerate the process of AS, in line with the previous studies. Therefore, we concluded that ox-LDL-induced hsa_circ_0003204 regulated endothelial cell injury through the miR-330-5p/Nod2 axis (Fig. 8).
Conclusions
In sum, hsa_circ_0003204 was reinforced in ox-LDL-induced HAECs. Moreover, hsa_circ_0003204 downregulation impeded the oxidative stress and apoptosis in ox-LDL-induced HAECs through inhibiting Nod2 via miR-330-5p. Also, we reported for the first time the role of hsa_circ_0003204 and the mechanism of the hsa_circ_0003204/miR-330-5p/Nod2 axis in AS progression. This study provided new insight for the prevention or treatment of AS.