22.11.2022
Redaktor:
Iwona Konarska
Źródło: Dermatology Review/Przegląd Dermatologiczny 3/2022 vol. 109
Zaburzenia immunologiczne w liszaju płaskim

Diagnostyka liszaja płaskiego na podstawie markerów immunologicznych jest nowym obszarem dermatologii. Ze względu na ograniczoną dotąd liczbę badań w tym zakresie nie zaleca się stosowania tej metody jako jedynego narzędzia diagnostycznego.
Aleksandra Ostrowska 1,2, Anna Susło 1,3
1Katedra i Klinika Dermatologiczna, Warszawski Uniwersytet Medyczny, Warszawa
2Szpital Wolski im. dr Anny Gostyńskiej, Warszawa
3Centralny Szpital Kliniczny MSWiA, Warszawa
Wprowadzenie
Liszaj płaski jest chorobą o podłożu immunologicznym, która powoduje przewlekły stan zapalny. Może zajmować skórę, błonę śluzową jamy ustnej, a także owłosioną skórę głowy, paznokcie i błony śluzowe narządów płciowych [1]. Wykwity w przebiegu liszaja płaskiego mają postać charakterystycznych wielobocznych grudek i blaszek o błyszczącej powierzchni, spoistych w badaniu palpacyjnym, którym towarzyszy świąd. Zmiany skórne są zwykle umiejscowione na powierzchniach zgięciowych nadgarstków, przedramion i kończyn dolnych [1]. Częstość występowania liszaja płaskiego w populacji wynosi od 0,4 proc. do 4 proc. Choroba może pojawić się w każdym wieku, ale najczęściej występuje między 30. a 60. rokiem życia, zwłaszcza u kobiet w okresie okołomenopauzalnym [1, 2].
Pomimo licznych badań etiologia liszaja płaskiego nadal jest nieznana [1–3]. Jako możliwe czynniki patogenetyczne wymienia się stosowanie leków (m.in. przeciwmalarycznych, sercowo-naczyniowych, soli złota, niesteroidowych leków przeciwzapalnych, hipoglikemizujących) oraz zakażenia wirusowe, zwłaszcza wirusem brodawczaka ludzkiego (human papillomavirus – HPV) i wirusem zapalenia wątroby typu C (hepatitis C virus – HCV).
Istnieją różne postacie morfologiczne liszaja płaskiego błony śluzowej jamy ustnej: nadżerkowa, zanikowa, pęcherzowa oraz najczęściej występująca siateczkowata [1]. Wśród cech histopatologicznych liszaja płaskiego należy wymienić zwyrodnienie wodniczkowe komórek warstwy podstawnej oraz obecność komórek zapalnych naciekających blaszkę właściwą błony śluzowej jamy ustnej. We wczesnych stadiach liszaja płaskiego przeważają makrofagi i limfocyty CD4+, natomiast w stadiach późniejszych stwierdza się głównie limfocyty T CD8+ [5]. Liszaj płaski ma nierzadko charakter przewlekły i oporny na leczenie lub przebiega z częstymi nawrotami pomimo prowadzonego leczenia [1]. U jednego pacjenta może współistnieć więcej niż jeden podtyp liszaja płaskiego [6].
Ze względu na obecność przewlekłego stanu zapalnego, który prowadzi do zmian mikrośrodowiska i uwolnienia wielu czynników powiązanych z powstawaniem, progresją i inwazją nowotworów, liszaj płaski jest uznawany za stan o potencjalnie zwiększonym ryzyku transformacji nowotworowej [7]. Ponadto uważa się, że w procesie onkogenezy związanej z liszajem płaskim mogą brać udział czynniki genetyczne, zaburzenia odżywiania, substancje toksyczne, mikroorganizmy oraz stan immunosupresji [4]. Największe ryzyko transformacji nowotworowej występuje w przypadku podtypu nadżerkowego [3]. Szacuje się, że prawdopodobieństwo procesu nowotworzenia związanego z liszajem płaskim w jamie ustnej wynosi ok. 1 proc. [8].
Rozpoznanie liszaja płaskiego na podstawie obrazu klinicznego często stanowi wyzwanie. Z tego względu istotne w diagnostyce jest badanie histologiczne [9]. Ostatnio wiele badań wskazuje na istnienie markerów immunologicznych liszaja płaskiego. Wiele z nich jest powszechnie obecnych w płynach ustrojowych. Daje to nadzieję na zwiększenie roli technik nieinwazyjnych w diagnostyce tej choroby. Poniżej przedstawiono zestawienie najnowszych markerów immunologicznych liszaja płaskiego.
Markery komórkowe
W przebiegu liszaja płaskiego obserwuje się wzmożoną ekspresję szeregu markerów, takich jak CD3+, CD4+, CD8+, CD19+ i CD83+, które są charakterystyczne dla limfocytów T i B [10]. Prawie 85 proc. komórek dominujących w zmianach chorobowych stanowią komórki CD27+. Są one obecne głęboko w blaszce właściwej, a nie w strefie podnabłonkowej, a zatem prawdopodobnie nie wpływają bezpośrednio na proces chorobowy, natomiast wykazują zależność z procesem autoimmunologicznym. Ekspresję CD38 w limfocytach B stwierdzono w 55 proc. zmian [11], a limfocyty T pamięci w 53,3 proc. zmian [12]. W badaniach wykazano również podwyższone stężenie limfocytów Th9. Limfocyty tego typu zwiększają nasilenie choroby wskutek wzrostu stężenia limfocytów Th17 lub pośrednio, poprzez synergistyczne działanie z Th17 [13]. Udział limfocytów Th1 i Th17 w zmianie chorobowej jest znamiennie większy niż w grupie kontrolnej. Wskazuje to, że limfocyty Th17 oraz wytwarzana przez nie cytokina Th17 mogą mieć istotne znaczenie w patogenezie liszaja płaskiego [14]. Wysoka liczba limfocytów CD8+ wiąże się natomiast z wysokim wskaźnikiem remisji [15].
Makrofagi są zwykle obecne w przewlekłych naciekach zapalnych przy komórkach podstawnych nabłonka. Jest to miejsce, w którym dochodzi do uszkodzenia komórek, a produkty makrofagów mogą wywoływać transformację złośliwą [16]. Po zakończeniu transformacji makrofagi M1 przechodzą w fenotyp M2 [17].
W naciekach występują także komórki tuczne [18]. Stwierdzono, że około 60% z nich to komórki tuczne zdegranulowane. W związku z tym one również mogą przyczyniać się do procesu transformacji złośliwej [16].
Zwiększona infiltracja komórek dendrytycznych CD11c+/CD123+ obserwowana jest głównie w naciekach z limfocytów T. Ich rola polega na przetwarzaniu i prezentacji antygenów keratynocytów, a także indukowaniu odporności limfocytów T [19]. Wykazano, że komórki dendrytyczne z czynnikiem XIIIa+ są rozmieszczone w nabłonku i głębokiej warstwie skóry właściwej. Poniżej nacieku z limfocytów T występowały komórki dendrytyczne CD34+, głównie w warstwie głębokiej skóry właściwej [20].
Liczba pomocniczych limfocytów T, makrofagów i komórek dendrytycznych jest znacząco wyższa we wczesnych stadiach liszaja płaskiego. Są to komórki prezentujące antygen, które wywołują odpowiedź zapalną. W zaawansowanym stadium choroby wzrasta liczba komórek obronnych, co prowadzi do apoptozy keratynocytów, które również mogą ulegać aktywacji. Następnie keratynocyty wydzielają chemokiny, które przyciągają limfocyty i inne komórki odpornościowe, aby wywołać proces transformacji w zmianach liszaja płaskiego [14].
Analiza stężeń białek wykazała wysokie stężenie receptora Toll-like 7 (TLR7), TLR8 i TLR9 [19]. Receptor TLR2 ulega dezaktywacji. Oznacza to, że nie dochodzi do pobudzenia szlaku jądrowego czynnika transkrypcyjnego NF-kB (NF-kB) i kinazy N-końcowej Jun (JNK)/p38 [10]. Niektóre badania wykazały, że stężenie TLR4 jest niskie w naskórku, a wysokie w skórze właściwej pacjentów, u których utrzymywał się nasilony naciek komórkowy [21]. Receptor ten indukuje ekspresję liganda receptora programowanej śmierci 1 (PD-L1) na keratynocytach i hamuje proliferację limfocytów T CD4+ oraz limfocytów T CD8+, jednocześnie indukując ich apoptozę [22].
Główną rolę w kolejnych szlakach sygnałowych odgrywa interferon α (IFN-α). Jest to potencjalny czynnik stymulujący rozwój choroby oraz występowanie zaostrzeń [19].
Niektórzy badacze wykryli przeciwciała przeciw komórkom podstawnym (anty-BCA) u 54% chorych z liszajem płaskim i potwierdzili, że utrzymują się w ich surowicy przez dłuższy czas. Być może dzieje się tak dlatego, że autoprzeciwciała są kierowane przeciwko zmienionym antygenom swoistym dla komórek podstawnych [23].
W innym badaniu stwierdzono wysokie stężenie liganda 10 chemokiny o motywie C-X-C (CXCL10) i interferonu γ (IFN-γ). Cytokiny te wpływają na przemieszczanie się limfocytów T do rozwijającej się zmiany [10].
Receptory antygenowe limfocytów T rozpoznają swoisty obcy antygen dzięki cząsteczkom głównego układu zgodności tkankowej (MHC) klasy I i II na keratynocytach podstawnych. Następnie limfocyty T CD4+ uwalniają interleukinę 2 (IL-2) i IFN-γ, aby aktywować limfocyty T CD8+ w celu wzbudzenia apoptozy keratynocytów przez czynnik martwicy nowotworów (TNF). Chemokina ta może być również uwalniana przez makrofagi i komórki tuczne. Pobudza wytwarzanie metaloproteinaz macierzy pozakomórkowej (MMPs), które aktywują apoptozę keratynocytów, wywołują zaburzenia w obrębie błon, a także sprzyjają procesowi kancerogenezy w nabłonku [16].
Za bodziec antygenowy w liszaju płaskim uznaje się białka szoku cieplnego. W przeprowadzonym badaniu stwierdzono podwyższone stężenie białka szoku cieplnego 27 (Hsp27) w keratynocytach oraz Hsp60 w keratynocytach i naciekających limfocytach. Stopień wybarwienia anty-Hsp90 wzrósł jedynie w błonie podstawnej dziąseł i podniebienia. Stężenie Hsp70 uległo natomiast zmniejszeniu [24].
Cytokiny
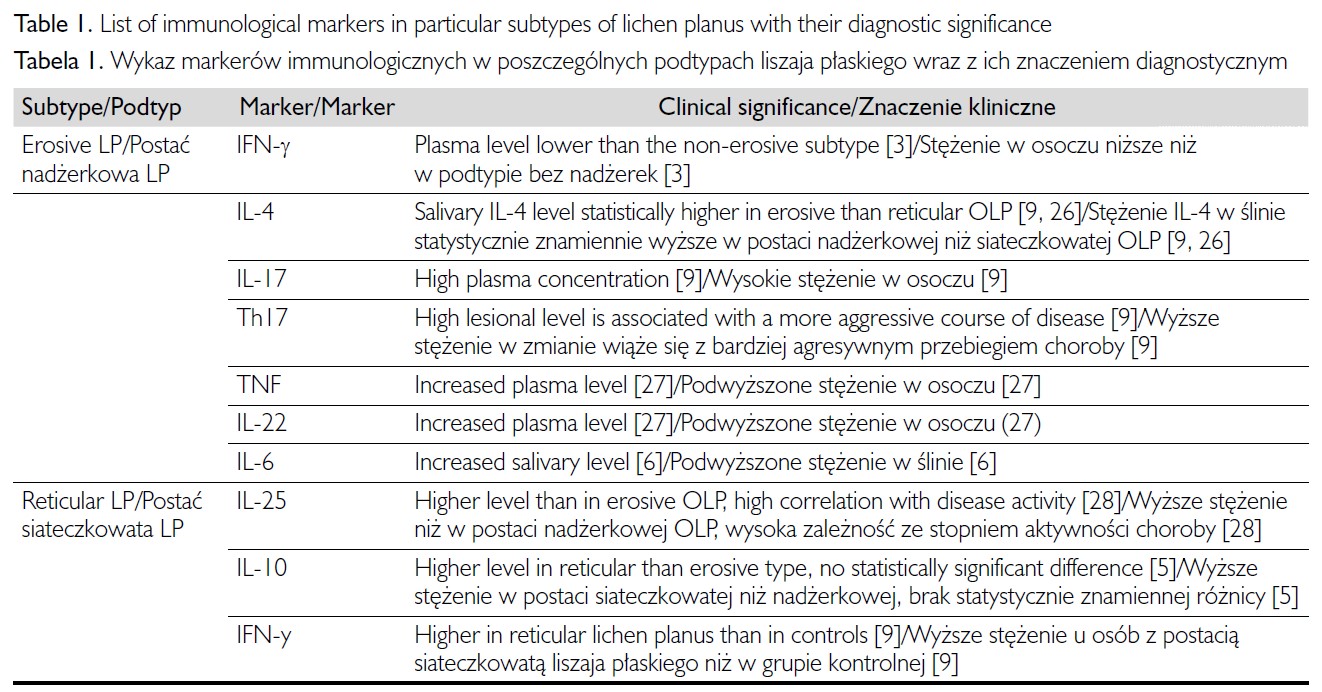
Główną rolę w nieinwazyjnej diagnostyce liszaja płaskiego odgrywają dziś interleukiny. Najczęściej ich stężenie oznaczane jest w osoczu lub ślinie pacjentów. Mają największe znaczenie w diagnostyce nadżerkowej i siateczkowatej postaci liszaja płaskiego, gdzie wykazano ich przydatność w ocenie stadium i stopnia aktywności choroby (tab. I).
Interleukiny
W przebiegu liszaja płaskiego obserwuje się podwyższone stężenie interleukin (IL) 1 (IL-1), IL-4, IL-6 i IL-8 [6, 8]. W dostępnym piśmiennictwie najwięcej danych dotyczy roli IL-6 jako potencjalnego markera w liszaju płaskim. IL-6 jest cytokiną prozapalną, która uczestniczy w procesach angiogenezy, wzrostu guza, inwazji i powstawania przerzutów [7]. Stwierdzono statystycznie znamiennie wyższe stężenia IL-6 w osoczu i ślinie pacjentów z liszajem płaskim w porównaniu z osobami zdrowymi [25]. Marker ten może być również przydatny w różnicowaniu postaci siateczkowatej od postaci nadżerkowej liszaja płaskiego, ponieważ wykazano, że jego stężenie w ślinie jest znamiennie wyższe u pacjentów z nadżerkową odmianą liszaja płaskiego niż z odmianą siateczkowatą [6]. Inne badania wskazują natomiast, że stężenie IL-6 w osoczu i w ślinie nie różni się statystycznie istotnie w powyższych podtypach liszaja płaskiego [7]. Podwyższone stężenie IL-6 jest również markerem prognostycznym w liszaju płaskim (powiązanym ze stopniem nasilenia i przebiegiem gojenia ran) i może być przydatne przy monitorowaniu odpowiedzi na leczenie [8].
IL-4 odgrywa ważną rolę w patogenezie liszaja płaskiego, zwłaszcza w mechanizmie Th2-zależnym, pobudzając wytwarzanie innych cytokin, syntezę przeciwciał i przebieg Th2-zależnej odpowiedzi humoralnej [26]. Stężenie tej interleukiny jest podwyższone w ślinie chorych z liszajem płaskim w porównaniu z grupą kontrolną [6, 8]. Wyniki większości badań wskazują, że stężenie IL-4 w osoczu lub ślinie jest wyższe w postaci nadżerkowej niż siateczkowatej liszaja płaskiego [9, 26–31]. Stężenie IL-4 w ślinie może również służyć jako biomarker ciężkości przebiegu choroby [5, 26]. W przeciwieństwie do IL-1, IL-6 i IL-8, IL-4 nie odgrywa istotnej roli w rozwoju raka kolczystokomórkowego (squamous cell carcinoma – SCC) na podłożu liszaja płaskiego [26].
Wyższe stężenie IL-25 i IL-4 wykryto w zmianach w przebiegu postaci siateczkowatej liszaja płaskiego w porównaniu z postacią nadżerkową. Ponadto stężenia IL-25 i IL-8 w ślinie wykazują zależność z nasileniem liszaja płaskiego [8, 28].
U pacjentów z liszajem płaskim obserwowano statystycznie znamiennie obniżone stężenie IL-2 oraz podwyższone stężenie IL-10 w surowicy w porównaniu z grupą kontrolną (p < 0,05) [5]. Stężenie IL-10 w surowicy było podwyższone zarówno u pacjentów z nadżerkową, jak i siateczkowatą postacią liszaja płaskiego, ale nie stwierdzono statystycznie znamiennych różnic w stężeniu tej interleukiny pomiędzy tymi dwiema grupami [5]. Wyniki te wskazują na dominację odpowiedzi Th2 u pacjentów z liszajem płaskim. Znaczenie podwyższonego stężenia IL-10 w diagnostyce liszaja płaskiego jest jednak niejasne, ponieważ uważa się, że wzrost stężenia tej interleukiny ma związek z odpowiedzią obronną gospodarza, której celem jest ochrona uszkodzonej tkanki przed dalszym urazem [32].
U chorych z liszajem płaskim odnotowano statystycznie znamienny wzrost średniego stężenia IL-18 w porównaniu z osobami zdrowymi z grupy kontrolnej [33]. Zaobserwowano ponadto, że w grupie pacjentów z postacią nadżerkową liszaja płaskiego (w porównaniu z postacią nienadżerkową) występuje podwyższone stężenie IL-18. Nie ma jednak pewności, czy różnica ta jest znamienna statystycznie i może być podstawą do rozpoznania podtypu nadżerkowego liszaja płaskiego [33, 34].
U pacjentów z liszajem płaskim obserwuje się zwiększoną ekspresję IL-17 w zmianach w obrębie jamy ustnej i na skórze oraz znamiennie podwyższone stężenie IL-17 w surowicy [9, 29]. Ponadto stężenie IL-17 w surowicy i ekspresja tej cytokiny w zmianach chorobowych mogą być pomocne w różnicowaniu podtypu nadżerkowego i siateczkowatego liszaja płaskiego [9]. Wysokie stężenie IL-17 w osoczu i zwiększoną ekspresję tej interleukiny obserwuje się u pacjentów z podtypem nadżerkowym liszaja płaskiego [9]. Ponadto wykazano zależność między ekspresją genu IL-17 a bardziej agresywnym przebiegiem choroby [9].
Na podstawie stężenia IL-22 w surowicy można również prognozować nawrót liszaja płaskiego: przy nawrocie postaci nadżerkowej stwierdzono 9-krotnie podwyższone stężenie IL-22 w porównaniu z grupą kontrolną [27]. Stężenie tej interleukiny może być również pomocnym narzędziem przy różnicowaniu przewlekłej postaci nadżerkowej i nienadżerkowej liszaja płaskiego. Podwyższone stężenie IL-22 wskazuje na postać nadżerkową, natomiast u chorych z przewlekłą postacią nienadżerkową stwierdza się statystycznie znamienne (p < 0,05) obniżenie stężenia IL-22 [27]. Zmniejszone stężenie IL-17 w skojarzeniu z obniżonym stężeniem IL-21 i IL-22 uznawane jest za marker remisji [35].
Chemokiny
Chemokiny są białkami prozapalnymi, które biorą udział w pobudzaniu leukocytów do napływu do miejsca zapalenia. Przeprowadzono liczne badania analizujące rolę chemokin w patogenezie liszaja płaskiego. Ich szczegółowa analiza wykracza jednak poza ramy niniejszego opracowania.
W tkankach stwierdzono znamiennie podwyższone stężenie CXCL9/10/11 i receptora 3 chemokiny o motywie C-X-C (CXCR3) [36, 37]. Ponadto u chorych z liszajem płaskim wykazano znamiennie wyższe stężenie CCL17 i CCR4 w surowicy, a także podwyższoną ekspresję CCL5-CCR5 w surowicy i tkankach, w porównaniu z grupą kontrolną [2, 38]. Wyniki są obiecujące, ponieważ otwierają nowe możliwości w zakresie opracowania leków dla chorych na liszaj płaski, ukierunkowanych na te osie terapeutyczne.
Inne cytokiny
TNF
Przez wiele lat badacze analizowali stężenie TNF w płynach ustrojowych w grupie pacjentów z liszajem płaskim. U tych chorych odnotowano statystycznie znamiennie podwyższone stężenie TNF w porównaniu z osobami zdrowymi z grupy kontrolnej [5]. Potwierdzono także zależność z podwyższonym stężeniem TNF w ślinie, co wskazuje, że może służyć jako marker progresji i początku choroby [8]. Ponadto w trzech badaniach wykazano związek między stężeniem TNF w ślinie a postacią kliniczną liszaja płaskiego: stężenie TNF w zmianach w postaci nadżerkowej było wyższe niż w postaci siateczkowatej. W niektórych badaniach zaobserwowano znamiennie podwyższone stężenie TNF w surowicy w porównaniu ze zdrowymi osobami z grupy kontrolnej [6, 39]. Podwyższone stężenie TNF w surowicy w przypadku liszaja płaskiego nadżerkowego może także służyć jako wskaźnik prognostyczny nawrotu choroby [27].
IFN-g
W badaniach obserwowano podwyższone stężenie IFN-g w ślinie oraz nadekspresję IFN-g w zmianach liszaja płaskiego w porównaniu z grupą kontrolną [6, 9]. W badaniu Liu i wsp. odnotowano niższe stężenia IFN-g zarówno w ślinie, jak i surowicy [31].
Stężenie IFN-g może być wykorzystywane do różnicowania podtypów liszaja płaskiego. Znamiennie niższe stężenie IFN-g w surowicy stwierdzano w podtypie nadżerkowym w porównaniu z postacią nienadżerkową choroby [40, 41]. Ponadto stężenie IFN-g w ślinie było wyższe u chorych z podtypem siateczkowatym niż u zdrowych osób z grupy kontrolnej [30]. Obserwacja ta wskazuje na udział Th1-zależnych reakcji zapalnych w procesie patogenezy.
Transformujący czynnik wzrostu β (TGF-β)
Rola TGF-β w zakresie kancerogenezy jest dwojaka: we wczesnych stadiach może hamować wzrost guza, natomiast w stadiach bardziej zaawansowanych może sprzyjać rozwojowi nowotworu poprzez indukcję angiogenezy i aktywację MMP-9 [4]. Na podstawie tych ustaleń oraz zgodnie ze stanowiskiem WHO, że liszaj płaski jest schorzeniem o potencjalnie złośliwym charakterze, przeprowadzono kilka badań, których celem była analiza ekspresji TGF-β. Stwierdzono znamiennie obniżone stężenie TGF-β w surowicy [39]. Ocena immunohistochemiczna TGF-β1 wykazała także umiarkowanie dodatnią ekspresję w nabłonku chorych z liszajem płaskim [42]. Obserwowano ponadto znamiennie zwiększoną ekspresję TGF-β u chorych z liszajem płaskim w porównaniu z prawidłową błoną śluzową jamy ustnej oraz dodatnią zależność pomiędzy ekspresją TGF-β1 a apoptozą komórek nabłonka [42]. Wyniki te wskazują na istotną rolę TGF-β w patogenezie liszaja płaskiego i rozwoju SCC na podłożu zmian liszaja płaskiego.
Inne markery
MMP 2, 9, 4, 7
Metaloproteinazy macierzy pozakomórkowej (MMPs) powodują uszkodzenia błony podstawnej i apoptozę keratynocytów [7]. Wysoki poziom ekspresji MMPs stwierdza się u pacjentów z liszajem płaskim [7]. Rubaci i wsp. wykazali podwyższoną ekspresję MMP-2 i MMP-7 w nabłonku i tkance łącznej u pacjentów z liszajem płaskim w porównaniu z grupą kontrolną [4]. Inni badacze obserwowali znamienne różnice w stężeniu MMPs pomiędzy postacią nadżerkową a nienadżerkową. Podwyższone stężenia MMP-1, MMP-2, MMP-3 i MMP-4 były obecne w podtypie nadżerkowym liszaja płaskiego [4]. W badaniu Wang i wsp. miejscowa ekspresja MMP-9 była podwyższona u pacjentów z liszajem płaskim w porównaniu z grupą kontrolną [13]. Inni badacze stwierdzili znamiennie podwyższone stężenie MMP-9 w tkankach przyzębia w tej grupie pacjentów [13].
Ze względu na destrukcyjne właściwości MMP badano zależność między ich ekspresją w zmianach chorobowych a procesem transformacji złośliwej. Na podstawie uzyskanych wyników ustalono, że MMP-2 i MMP-9 stanowią potencjalne markery złośliwej transformacji liszaja płaskiego [4, 7].
MHC
W zmianach wywoływanych przez liszaj płaski stwierdzano nadekspresję MHC klasy I i II. Ponadto obserwowano, że blokowanie wyłącznie HLA-DR, w przeciwieństwie do HLA-DP i DQ, chroni przed śmiercią keratynocytów [43]. W przypadku blokowania MHC klasy I odnotowywano bardziej nasilone oddziaływanie hamujące na śmierć keratynocytów indukowaną przez limfocyty T, co wskazuje na udział MHC klasy I w prezentacji antygenu w liszaju płaskim [43].
Cząsteczka adhezji komórkowej będąca adresyną błon śluzowych 1 (MAdCAM-1)
Stężenie MAdCAM-1 było w sposób znamienny obniżone w subpopulacji pacjentów z liszajem płaskim. Stwierdzono, że MAdCAM-1 może mieć znaczenie jako wskaźnik stanu zapalnego, ale nie jako marker aktywności choroby [44].
Wnioski
W liszaju płaskim nie ustalono dotąd czynnika patogenetycznego odpowiadającego za rozwój tej choroby, jednak patogeneza schorzenia jest ściśle związana z zaburzeniami immunologicznymi. Nieprawidłowości immunologiczne obejmują zróżnicowaną populację komórek odpornościowych, co może wskazywać na istnienie wspólnej cząsteczki inicjującej ten proces. Interesująca jest obserwacja, że liczba komórek prezentujących antygen wzrasta w stadiach poprzedzających rozwój pełnoobjawowej choroby.
Znajomość odchyleń w stężeniach markerów immunologicznych, m.in. cytokiny w ślinie i surowicy, może być przydatna w precyzyjnej diagnostyce bez użycia metod inwazyjnych, takich jak biopsja. W postaci siateczkowatej liszaja płaskiego obserwowane jest podwyższone stężenie IL-25, IL-10 i IFN-g, natomiast w postaci nadżerkowej występuje zwiększone stężenie IL-4, IL-6, IL-6, IL-17, IL-22 i TNF. Analiza stężenia IL-6 w ślinie może być przydatna w ocenie stopnia zaawansowania choroby i monitorowaniu odpowiedzi na leczenie. Podwyższone stężenie IL-22 w surowicy może służyć jako marker nawrotu choroby. W przypadku remisji obserwowane są obniżone stężenia IL-17, IL-21 i IL-22.
Należy zawsze pamiętać, że z powodu przewlekłego stanu zapalnego liszaj płaski jest stanem przednowotworowym. Wyniki, takie jak podwyższone stężenie MMP-2 i MMP-9 oraz zmniejszone stężenie TGF-β, sygnalizują wzrost potencjalnego ryzyka transformacji nowotworowej.
Przedstawione powyżej ustalenia mogą być przydatne w opracowaniu skutecznego leczenia przyczynowego, szczególnie w przypadkach opornych na standardową terapię. Diagnostyka liszaja płaskiego na podstawie markerów immunologicznych jest nowym obszarem dermatologii. Ze względu na ograniczoną dotąd liczbę badań w tym zakresie nie zaleca się stosowania tej metody jako jedynego narzędzia diagnostycznego. Potrzebne są dalsze badania i obserwacje, a pomimo obiecujących wyników nie można na razie wyeliminować złotego standardu diagnostycznego – badania histopatologicznego.
Konflikt interesów
Autorki nie zgłaszają konfliktu interesów.
Piśmiennictwo
1. Usatine R.P., Tinitigan M.: Diagnosis and treatment of lichen planus. Am Fam Physician 2011, 84, 53-60.
2. Shan J., Shen C., Fang J., Li S., Fan Y.: Potential roles of the CCL17-CCR4 axis in the immunopathogenesis of oral lichen planus. J Oral Pathol Med 2020, 49, 328-334.
3. Mozaffari H.R., Sharifi R., Hayati M., Imani M.M., Lopez-Jornet P., Golshah A., et al.: Evaluation of serum and salivary interferon-γ levels in patients with oral lichen planus: a systematic review and meta-analysis of case-control studies. Oral Surg Oral Med Oral Pathol Oral Radiol 2019, 127, 210-217.
4. Payeras M.R., Cherubini K., Figueiredo M.A., Salum F.G.: Oral lichen planus: focus on etiopathogenesis. Arch Oral Biol 2013, 58, 1057-1069.
5. Pekiner F.N., Demirel G.Y., Borahan M.O., Ozbayrak S.: Cytokine profiles in serum of patients with oral lichen planus. Cytokine 2012, 60, 701-706.
6. Humberto J.S.M., Pavanin J.V., Rocha M., Motta A.C.F.: Cytokines, cortisol, and nitric oxide as salivary biomarkers in oral lichen planus: a systematic review. Braz Oral Res 2018, 32, e82.
7. Mignogna M.D., Fedele S., Lo Russo L., Lo Muzio L., Bucci E.: Immune activation and chronic inflammation as the cause of malignancy in oral lichen planus: is there any evidence? Oral Oncol 2004, 40, 120-130.
8. Melguizo-Rodriguez L., Costela-Ruiz V.J., Manzano-Moreno F.J., Ruiz C., Illescas-Montes R.: Salivary biomarkers and their application in the diagnosis and monitoring of the most common oral pathologies. Int J Mol Sci 2020, 21, 5173.
9. Carvalho M., Cavalieri D., Do Nascimento S., Lourenço T.G.B., Ramos D.V.R., Pasqualin D.D.C., et al.: Cytokines levels and salivary microbiome play a potential role in oral lichen planus diagnosis. Sci Rep 2019, 9, 18137.
10. Sinon S.H., Rich A.M., Firth N.A., Seymour G.J.: Does the toll-like receptor 2 (TLR2) play a role in oral mucosal lichen planus (OMLP)? Pathology 2011, 43, S65.
11. Mattila R., Ahlfors E., Syrjänen S.: CD27 and CD38 lymphocytes are detected in oral lichen planus lesions. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2011, 111, 211-217.
12. Devi M., Vijayalakshmi D., Dhivya K., Janane M.: Memory T cells (CD45RO) role and evaluation in pathogenesis of lichen planus and lichenoid mucositis. J Clin Diagn Res 2017, 11, ZC84-ZC6.
13. Wang H., Guan X., Luo Z., Liu Y., Ren Q., Zhao X.: The association and potentially destructive role of Th9/IL-9 is synergistic with Th17 cells by elevating MMP9 production in local lesions of oral lichen planus. J Oral Pathol Med 2018, 47, 425-433.
14. Payeras M.R., Cherubini K., Figueiredo M.A., Salum F.G.: Oral lichen planus: focus on etiopathogenesis. Arch Oral Biol 2013, 58, 1057-1069.
15. Enomoto A., Sato E., Yasuda T., Isomura T., Nagao T., Chikazu D.: Intraepithelial CD8+ lymphocytes as a predictive diagnostic biomarker for the remission of oral lichen planus. Human Pathol 2018, 74, 43-53.
16. Mignogna M.D., Fedele S., Lo Russo L., Lo Muzio L., Bucci E.: Immune activation and chronic inflammation as the cause of malignancy in oral lichen planus: is there any evidence ? Oral Oncol 2004, 40, 120-130.
17. Jablonska E., Garley M., Surazynski A., Grubczak K., Iwaniuk A., Borys J., et al.: Neutrophil extracellular traps (NETs) formation induced by TGF-beta in oral lichen planus – possible implications for the development of oral cancer. Immunobiology 2020, 225, 151901.
18. Ramalingam S., Malathi N., Thamizhchelvan H., Sangeetha N., Rajan S.T.: Role of mast cells in oral lichen planus and oral lichenoid reactions. Autoimm Dis 2018, 2018, 7936564.
19. Wang Y., Shang S., Sun Q., Chen J., Du G., Nie H., et al.: Increased infiltration of CD11 c(+)/CD123(+) dendritic cell subsets and upregulation of TLR/IFN-α signaling participate in pathogenesis of oral lichen planus. Oral Surg Oral Med Oral Pathol Oral Radiol 2018, 125, 459-467.e2.
20. Cardozo A.L., Moura-Castro C., Figueiredo M., Cuzzi T., Ramos-e-Silva M.: Oral lichen planus and dermal dendrocytes. Actas Dermosifiliogr 2009, 100, 46-52.
21. de Carvalho G.C., Hirata F.Y.A., Domingues R., Figueiredo C.A., Zaniboni M.C., Pereira N.V., et al.: Up-regulation of HMGB1 and TLR4 in skin lesions of lichen planus. Arch Dermatol Res 2018, 310, 523-528.
22. Zhang J., Tan Y.Q., Wei M.H., Ye X.J., Chen G.Y., Lu R., et al.: TLR4-induced B7-H1 on keratinocytes negatively regulates CD4(+) T cells and CD8(+) T cells responses in oral lichen planus. Exp Dermatol 2017, 26, 409-415.
23. Lin S.C., Sun A., Wu Y.C., Chiang C.P.: Presence of anti-basal cell antibodies in oral lichen planus. J Am Acad Dermatol 1992, 26, 943-947.
24. Bramanti T.E., Dekker N.P., Lozada-Nur F., Sauk J.J., Regezi J.A.: Heat shock (stress) proteins and γδ T lymphocytes in oral lichen planus. Oral Surg Oral Med Oral Pathol Oral Radiol Endodontol 1995, 80, 698-704.
25. Liu J., Shi Q., Yang S., Wang Q., Xu J., Guo B.: The relationship between levels of salivary and serum interleukin-6 and oral lichen planus: a systematic review and meta-analysis. J Am Dental Assoc 2017, 148, 743-749.e9.
26. Mehrbani S.P., Motahari P., Azar F.P., Ahari M.A.: Role of interleukin-4 in pathogenesis of oral lichen planus: a systematic review. Med Oral Patol Oral Cirugia Bucal 2020, 25, e410-e415.
27. Kurchenko A., Drannik G., Rehuretska R., duBuske L.: Assessment of select serum cytokines TNF-a and IL-22 in oral lichen planus patients. Ann Allergy Asthma Immunol 2018, 121, 5 Suppl, S50.
28. Wang H., Jiang Y., Wang H., Luo Z., Wang Y., Guan X.: IL-25 promotes Th2-type reactions and correlates with disease severity in the pathogenesis of oral lichen planus. Arch Oral Biol 2019, 98, 115-121.
29. Zychowska M., Batycka-Baran A., Baran W.: Increased serum level and high tissue immunoexpression of interleukin 17 in cutaneous lichen planus: a novel therapeutic target for recalcitrant cases? Dis Markers 2020, 2020, 6521274.
30. Mozaffari H.R., Zavattaro E., Saeedi M., Lopez-Jornet P., Sadeghi M., Safaei M., et al.: Serum and salivary interleukin-4 levels in patients with oral lichen planus: a systematic review and meta-analysis. Oral Surg Oral Med Oral Pathol Oral Radiol 2019, 128, 123-131.
31. Liu W.Z., He M.J., Long L., Mu D.L., Xu M.S., Xing X., et al.: Interferon-γ and interleukin-4 detected in serum and saliva from patients with oral lichen planus. Int J Oral Sci 2014, 6, 22-26.
32. Dan H., Liu W., Wang J., Wang Z., Wu R., Chen Q., et al.: Elevated IL-10 concentrations in serum and saliva from patients with oral lichen planus. Quintessence Int 2011, 42, 157-163.
33. Negi D., Urs A.B., Kumar P., Mahajan B., Singh H., Polipalli S.K., et al.: Assessment of interleukin-18 gene polymorphism and serum levels in oral lichen planus in an Indian population. J Oral Pathol Med 2019, 48, 244-250.
34. Zhang Y., Liu W., Zhang S., Dan H., Lu R., Wang F., et al.: Salivary and serum interleukin-18 in patients with oral lichen planus: a study in an ethnic Chinese population. Inflammation 2012, 35, 399-404.
35. Drannik G.N., Kurchenko A.I., Rehuretska R.A., Drannik A.G., DuBuske L.M: Th-17 cytokines in oral lichen planus. J Allergy Clin Immunol 2014, 133, AB186.
36. Marshall A., Celentano A., Cirillo N., McCullough M., Porter S.: Tissue-specific regulation of CXCL9/10/11 chemokines in keratinocytes: implications for oral inflammatory disease. PLoS One 2017, 12, e0172821.
37. Fang J., Wang C., Shen C., Shan J., Wang X., Liu L., et al.: The expression of CXCL10/CXCR3 and effect of the axis on the function of T lymphocyte involved in oral lichen planus. Inflammation 2019, 42, 799-810.
38. Shan J., Li S., Wang C., Liu L., Wang X., Zhu D., et al.: Expression and biological functions of the CCL5-CCR5 axis in oral lichen planus. Exp Dermatol 2019, 28, 816-821.
39. Taghavi Zenouz A., Pouralibaba F., Babaloo Z., Mehdipour M., Jamali Z.: Evaluation of serum TNF-α and TGF-β in patients with oral lichen planus. J Dent Res Dent Clin Dent Prospects 2012, 6, 143-147.
40. Ding M., Zeng J., Sroussi H., Yu J., Xu J., Cheng X., et al.: Interactions between Golli-MBP and Th1/Th2 cytokines in patients with oral lichen planus. Oral Dis 2014, 20, 205-211.
41. Xie S., Ding L., Xiong Z., Zhu S.: Implications of Th1 and Th17 cells in pathogenesis of oral lichen planus. J Huazhong Univ Sci Technolog Med Sci 2012, 32, 451-457.
42. Chen Z.L., Deng G.H., Chen H.B., Cheng J.: Expression of TGF-beta1, Smad7 and cell apoptosis in epithelium of oral lichen planus. Zhonghua Kou Qiang Yi Xue Za Zhi 2008, 43, 95-98.
43. Shao S., Tsoi L.C., Sarkar M.K., Xing X., Xue K., Uppala R., et al.: IFN-γ enhances cell-mediated cytotoxicity against keratinocytes via JAK2/STAT1 in lichen planus. Sci Transl Med 2019, 11, eaav7561.
44. Baran A., Nowowiejska J., Kamiński T.W., Krahel J.A., Flisiak I.: Circulating MAdCAM-1 and ITGB7 in patients with plaque psoriasis and eruptive lichen planus-preliminary data. Biology 2021, 10, 1129.
1Katedra i Klinika Dermatologiczna, Warszawski Uniwersytet Medyczny, Warszawa
2Szpital Wolski im. dr Anny Gostyńskiej, Warszawa
3Centralny Szpital Kliniczny MSWiA, Warszawa
Wprowadzenie
Liszaj płaski jest chorobą o podłożu immunologicznym, która powoduje przewlekły stan zapalny. Może zajmować skórę, błonę śluzową jamy ustnej, a także owłosioną skórę głowy, paznokcie i błony śluzowe narządów płciowych [1]. Wykwity w przebiegu liszaja płaskiego mają postać charakterystycznych wielobocznych grudek i blaszek o błyszczącej powierzchni, spoistych w badaniu palpacyjnym, którym towarzyszy świąd. Zmiany skórne są zwykle umiejscowione na powierzchniach zgięciowych nadgarstków, przedramion i kończyn dolnych [1]. Częstość występowania liszaja płaskiego w populacji wynosi od 0,4 proc. do 4 proc. Choroba może pojawić się w każdym wieku, ale najczęściej występuje między 30. a 60. rokiem życia, zwłaszcza u kobiet w okresie okołomenopauzalnym [1, 2].
Pomimo licznych badań etiologia liszaja płaskiego nadal jest nieznana [1–3]. Jako możliwe czynniki patogenetyczne wymienia się stosowanie leków (m.in. przeciwmalarycznych, sercowo-naczyniowych, soli złota, niesteroidowych leków przeciwzapalnych, hipoglikemizujących) oraz zakażenia wirusowe, zwłaszcza wirusem brodawczaka ludzkiego (human papillomavirus – HPV) i wirusem zapalenia wątroby typu C (hepatitis C virus – HCV).
Istnieją różne postacie morfologiczne liszaja płaskiego błony śluzowej jamy ustnej: nadżerkowa, zanikowa, pęcherzowa oraz najczęściej występująca siateczkowata [1]. Wśród cech histopatologicznych liszaja płaskiego należy wymienić zwyrodnienie wodniczkowe komórek warstwy podstawnej oraz obecność komórek zapalnych naciekających blaszkę właściwą błony śluzowej jamy ustnej. We wczesnych stadiach liszaja płaskiego przeważają makrofagi i limfocyty CD4+, natomiast w stadiach późniejszych stwierdza się głównie limfocyty T CD8+ [5]. Liszaj płaski ma nierzadko charakter przewlekły i oporny na leczenie lub przebiega z częstymi nawrotami pomimo prowadzonego leczenia [1]. U jednego pacjenta może współistnieć więcej niż jeden podtyp liszaja płaskiego [6].
Ze względu na obecność przewlekłego stanu zapalnego, który prowadzi do zmian mikrośrodowiska i uwolnienia wielu czynników powiązanych z powstawaniem, progresją i inwazją nowotworów, liszaj płaski jest uznawany za stan o potencjalnie zwiększonym ryzyku transformacji nowotworowej [7]. Ponadto uważa się, że w procesie onkogenezy związanej z liszajem płaskim mogą brać udział czynniki genetyczne, zaburzenia odżywiania, substancje toksyczne, mikroorganizmy oraz stan immunosupresji [4]. Największe ryzyko transformacji nowotworowej występuje w przypadku podtypu nadżerkowego [3]. Szacuje się, że prawdopodobieństwo procesu nowotworzenia związanego z liszajem płaskim w jamie ustnej wynosi ok. 1 proc. [8].
Rozpoznanie liszaja płaskiego na podstawie obrazu klinicznego często stanowi wyzwanie. Z tego względu istotne w diagnostyce jest badanie histologiczne [9]. Ostatnio wiele badań wskazuje na istnienie markerów immunologicznych liszaja płaskiego. Wiele z nich jest powszechnie obecnych w płynach ustrojowych. Daje to nadzieję na zwiększenie roli technik nieinwazyjnych w diagnostyce tej choroby. Poniżej przedstawiono zestawienie najnowszych markerów immunologicznych liszaja płaskiego.
Markery komórkowe
W przebiegu liszaja płaskiego obserwuje się wzmożoną ekspresję szeregu markerów, takich jak CD3+, CD4+, CD8+, CD19+ i CD83+, które są charakterystyczne dla limfocytów T i B [10]. Prawie 85 proc. komórek dominujących w zmianach chorobowych stanowią komórki CD27+. Są one obecne głęboko w blaszce właściwej, a nie w strefie podnabłonkowej, a zatem prawdopodobnie nie wpływają bezpośrednio na proces chorobowy, natomiast wykazują zależność z procesem autoimmunologicznym. Ekspresję CD38 w limfocytach B stwierdzono w 55 proc. zmian [11], a limfocyty T pamięci w 53,3 proc. zmian [12]. W badaniach wykazano również podwyższone stężenie limfocytów Th9. Limfocyty tego typu zwiększają nasilenie choroby wskutek wzrostu stężenia limfocytów Th17 lub pośrednio, poprzez synergistyczne działanie z Th17 [13]. Udział limfocytów Th1 i Th17 w zmianie chorobowej jest znamiennie większy niż w grupie kontrolnej. Wskazuje to, że limfocyty Th17 oraz wytwarzana przez nie cytokina Th17 mogą mieć istotne znaczenie w patogenezie liszaja płaskiego [14]. Wysoka liczba limfocytów CD8+ wiąże się natomiast z wysokim wskaźnikiem remisji [15].
Makrofagi są zwykle obecne w przewlekłych naciekach zapalnych przy komórkach podstawnych nabłonka. Jest to miejsce, w którym dochodzi do uszkodzenia komórek, a produkty makrofagów mogą wywoływać transformację złośliwą [16]. Po zakończeniu transformacji makrofagi M1 przechodzą w fenotyp M2 [17].
W naciekach występują także komórki tuczne [18]. Stwierdzono, że około 60% z nich to komórki tuczne zdegranulowane. W związku z tym one również mogą przyczyniać się do procesu transformacji złośliwej [16].
Zwiększona infiltracja komórek dendrytycznych CD11c+/CD123+ obserwowana jest głównie w naciekach z limfocytów T. Ich rola polega na przetwarzaniu i prezentacji antygenów keratynocytów, a także indukowaniu odporności limfocytów T [19]. Wykazano, że komórki dendrytyczne z czynnikiem XIIIa+ są rozmieszczone w nabłonku i głębokiej warstwie skóry właściwej. Poniżej nacieku z limfocytów T występowały komórki dendrytyczne CD34+, głównie w warstwie głębokiej skóry właściwej [20].
Liczba pomocniczych limfocytów T, makrofagów i komórek dendrytycznych jest znacząco wyższa we wczesnych stadiach liszaja płaskiego. Są to komórki prezentujące antygen, które wywołują odpowiedź zapalną. W zaawansowanym stadium choroby wzrasta liczba komórek obronnych, co prowadzi do apoptozy keratynocytów, które również mogą ulegać aktywacji. Następnie keratynocyty wydzielają chemokiny, które przyciągają limfocyty i inne komórki odpornościowe, aby wywołać proces transformacji w zmianach liszaja płaskiego [14].
Analiza stężeń białek wykazała wysokie stężenie receptora Toll-like 7 (TLR7), TLR8 i TLR9 [19]. Receptor TLR2 ulega dezaktywacji. Oznacza to, że nie dochodzi do pobudzenia szlaku jądrowego czynnika transkrypcyjnego NF-kB (NF-kB) i kinazy N-końcowej Jun (JNK)/p38 [10]. Niektóre badania wykazały, że stężenie TLR4 jest niskie w naskórku, a wysokie w skórze właściwej pacjentów, u których utrzymywał się nasilony naciek komórkowy [21]. Receptor ten indukuje ekspresję liganda receptora programowanej śmierci 1 (PD-L1) na keratynocytach i hamuje proliferację limfocytów T CD4+ oraz limfocytów T CD8+, jednocześnie indukując ich apoptozę [22].
Główną rolę w kolejnych szlakach sygnałowych odgrywa interferon α (IFN-α). Jest to potencjalny czynnik stymulujący rozwój choroby oraz występowanie zaostrzeń [19].
Niektórzy badacze wykryli przeciwciała przeciw komórkom podstawnym (anty-BCA) u 54% chorych z liszajem płaskim i potwierdzili, że utrzymują się w ich surowicy przez dłuższy czas. Być może dzieje się tak dlatego, że autoprzeciwciała są kierowane przeciwko zmienionym antygenom swoistym dla komórek podstawnych [23].
W innym badaniu stwierdzono wysokie stężenie liganda 10 chemokiny o motywie C-X-C (CXCL10) i interferonu γ (IFN-γ). Cytokiny te wpływają na przemieszczanie się limfocytów T do rozwijającej się zmiany [10].
Receptory antygenowe limfocytów T rozpoznają swoisty obcy antygen dzięki cząsteczkom głównego układu zgodności tkankowej (MHC) klasy I i II na keratynocytach podstawnych. Następnie limfocyty T CD4+ uwalniają interleukinę 2 (IL-2) i IFN-γ, aby aktywować limfocyty T CD8+ w celu wzbudzenia apoptozy keratynocytów przez czynnik martwicy nowotworów (TNF). Chemokina ta może być również uwalniana przez makrofagi i komórki tuczne. Pobudza wytwarzanie metaloproteinaz macierzy pozakomórkowej (MMPs), które aktywują apoptozę keratynocytów, wywołują zaburzenia w obrębie błon, a także sprzyjają procesowi kancerogenezy w nabłonku [16].
Za bodziec antygenowy w liszaju płaskim uznaje się białka szoku cieplnego. W przeprowadzonym badaniu stwierdzono podwyższone stężenie białka szoku cieplnego 27 (Hsp27) w keratynocytach oraz Hsp60 w keratynocytach i naciekających limfocytach. Stopień wybarwienia anty-Hsp90 wzrósł jedynie w błonie podstawnej dziąseł i podniebienia. Stężenie Hsp70 uległo natomiast zmniejszeniu [24].
Cytokiny
Główną rolę w nieinwazyjnej diagnostyce liszaja płaskiego odgrywają dziś interleukiny. Najczęściej ich stężenie oznaczane jest w osoczu lub ślinie pacjentów. Mają największe znaczenie w diagnostyce nadżerkowej i siateczkowatej postaci liszaja płaskiego, gdzie wykazano ich przydatność w ocenie stadium i stopnia aktywności choroby (tab. I).
Interleukiny
W przebiegu liszaja płaskiego obserwuje się podwyższone stężenie interleukin (IL) 1 (IL-1), IL-4, IL-6 i IL-8 [6, 8]. W dostępnym piśmiennictwie najwięcej danych dotyczy roli IL-6 jako potencjalnego markera w liszaju płaskim. IL-6 jest cytokiną prozapalną, która uczestniczy w procesach angiogenezy, wzrostu guza, inwazji i powstawania przerzutów [7]. Stwierdzono statystycznie znamiennie wyższe stężenia IL-6 w osoczu i ślinie pacjentów z liszajem płaskim w porównaniu z osobami zdrowymi [25]. Marker ten może być również przydatny w różnicowaniu postaci siateczkowatej od postaci nadżerkowej liszaja płaskiego, ponieważ wykazano, że jego stężenie w ślinie jest znamiennie wyższe u pacjentów z nadżerkową odmianą liszaja płaskiego niż z odmianą siateczkowatą [6]. Inne badania wskazują natomiast, że stężenie IL-6 w osoczu i w ślinie nie różni się statystycznie istotnie w powyższych podtypach liszaja płaskiego [7]. Podwyższone stężenie IL-6 jest również markerem prognostycznym w liszaju płaskim (powiązanym ze stopniem nasilenia i przebiegiem gojenia ran) i może być przydatne przy monitorowaniu odpowiedzi na leczenie [8].
IL-4 odgrywa ważną rolę w patogenezie liszaja płaskiego, zwłaszcza w mechanizmie Th2-zależnym, pobudzając wytwarzanie innych cytokin, syntezę przeciwciał i przebieg Th2-zależnej odpowiedzi humoralnej [26]. Stężenie tej interleukiny jest podwyższone w ślinie chorych z liszajem płaskim w porównaniu z grupą kontrolną [6, 8]. Wyniki większości badań wskazują, że stężenie IL-4 w osoczu lub ślinie jest wyższe w postaci nadżerkowej niż siateczkowatej liszaja płaskiego [9, 26–31]. Stężenie IL-4 w ślinie może również służyć jako biomarker ciężkości przebiegu choroby [5, 26]. W przeciwieństwie do IL-1, IL-6 i IL-8, IL-4 nie odgrywa istotnej roli w rozwoju raka kolczystokomórkowego (squamous cell carcinoma – SCC) na podłożu liszaja płaskiego [26].
Wyższe stężenie IL-25 i IL-4 wykryto w zmianach w przebiegu postaci siateczkowatej liszaja płaskiego w porównaniu z postacią nadżerkową. Ponadto stężenia IL-25 i IL-8 w ślinie wykazują zależność z nasileniem liszaja płaskiego [8, 28].
U pacjentów z liszajem płaskim obserwowano statystycznie znamiennie obniżone stężenie IL-2 oraz podwyższone stężenie IL-10 w surowicy w porównaniu z grupą kontrolną (p < 0,05) [5]. Stężenie IL-10 w surowicy było podwyższone zarówno u pacjentów z nadżerkową, jak i siateczkowatą postacią liszaja płaskiego, ale nie stwierdzono statystycznie znamiennych różnic w stężeniu tej interleukiny pomiędzy tymi dwiema grupami [5]. Wyniki te wskazują na dominację odpowiedzi Th2 u pacjentów z liszajem płaskim. Znaczenie podwyższonego stężenia IL-10 w diagnostyce liszaja płaskiego jest jednak niejasne, ponieważ uważa się, że wzrost stężenia tej interleukiny ma związek z odpowiedzią obronną gospodarza, której celem jest ochrona uszkodzonej tkanki przed dalszym urazem [32].
U chorych z liszajem płaskim odnotowano statystycznie znamienny wzrost średniego stężenia IL-18 w porównaniu z osobami zdrowymi z grupy kontrolnej [33]. Zaobserwowano ponadto, że w grupie pacjentów z postacią nadżerkową liszaja płaskiego (w porównaniu z postacią nienadżerkową) występuje podwyższone stężenie IL-18. Nie ma jednak pewności, czy różnica ta jest znamienna statystycznie i może być podstawą do rozpoznania podtypu nadżerkowego liszaja płaskiego [33, 34].
U pacjentów z liszajem płaskim obserwuje się zwiększoną ekspresję IL-17 w zmianach w obrębie jamy ustnej i na skórze oraz znamiennie podwyższone stężenie IL-17 w surowicy [9, 29]. Ponadto stężenie IL-17 w surowicy i ekspresja tej cytokiny w zmianach chorobowych mogą być pomocne w różnicowaniu podtypu nadżerkowego i siateczkowatego liszaja płaskiego [9]. Wysokie stężenie IL-17 w osoczu i zwiększoną ekspresję tej interleukiny obserwuje się u pacjentów z podtypem nadżerkowym liszaja płaskiego [9]. Ponadto wykazano zależność między ekspresją genu IL-17 a bardziej agresywnym przebiegiem choroby [9].
Na podstawie stężenia IL-22 w surowicy można również prognozować nawrót liszaja płaskiego: przy nawrocie postaci nadżerkowej stwierdzono 9-krotnie podwyższone stężenie IL-22 w porównaniu z grupą kontrolną [27]. Stężenie tej interleukiny może być również pomocnym narzędziem przy różnicowaniu przewlekłej postaci nadżerkowej i nienadżerkowej liszaja płaskiego. Podwyższone stężenie IL-22 wskazuje na postać nadżerkową, natomiast u chorych z przewlekłą postacią nienadżerkową stwierdza się statystycznie znamienne (p < 0,05) obniżenie stężenia IL-22 [27]. Zmniejszone stężenie IL-17 w skojarzeniu z obniżonym stężeniem IL-21 i IL-22 uznawane jest za marker remisji [35].
Chemokiny
Chemokiny są białkami prozapalnymi, które biorą udział w pobudzaniu leukocytów do napływu do miejsca zapalenia. Przeprowadzono liczne badania analizujące rolę chemokin w patogenezie liszaja płaskiego. Ich szczegółowa analiza wykracza jednak poza ramy niniejszego opracowania.
W tkankach stwierdzono znamiennie podwyższone stężenie CXCL9/10/11 i receptora 3 chemokiny o motywie C-X-C (CXCR3) [36, 37]. Ponadto u chorych z liszajem płaskim wykazano znamiennie wyższe stężenie CCL17 i CCR4 w surowicy, a także podwyższoną ekspresję CCL5-CCR5 w surowicy i tkankach, w porównaniu z grupą kontrolną [2, 38]. Wyniki są obiecujące, ponieważ otwierają nowe możliwości w zakresie opracowania leków dla chorych na liszaj płaski, ukierunkowanych na te osie terapeutyczne.
Inne cytokiny
TNF
Przez wiele lat badacze analizowali stężenie TNF w płynach ustrojowych w grupie pacjentów z liszajem płaskim. U tych chorych odnotowano statystycznie znamiennie podwyższone stężenie TNF w porównaniu z osobami zdrowymi z grupy kontrolnej [5]. Potwierdzono także zależność z podwyższonym stężeniem TNF w ślinie, co wskazuje, że może służyć jako marker progresji i początku choroby [8]. Ponadto w trzech badaniach wykazano związek między stężeniem TNF w ślinie a postacią kliniczną liszaja płaskiego: stężenie TNF w zmianach w postaci nadżerkowej było wyższe niż w postaci siateczkowatej. W niektórych badaniach zaobserwowano znamiennie podwyższone stężenie TNF w surowicy w porównaniu ze zdrowymi osobami z grupy kontrolnej [6, 39]. Podwyższone stężenie TNF w surowicy w przypadku liszaja płaskiego nadżerkowego może także służyć jako wskaźnik prognostyczny nawrotu choroby [27].
IFN-g
W badaniach obserwowano podwyższone stężenie IFN-g w ślinie oraz nadekspresję IFN-g w zmianach liszaja płaskiego w porównaniu z grupą kontrolną [6, 9]. W badaniu Liu i wsp. odnotowano niższe stężenia IFN-g zarówno w ślinie, jak i surowicy [31].
Stężenie IFN-g może być wykorzystywane do różnicowania podtypów liszaja płaskiego. Znamiennie niższe stężenie IFN-g w surowicy stwierdzano w podtypie nadżerkowym w porównaniu z postacią nienadżerkową choroby [40, 41]. Ponadto stężenie IFN-g w ślinie było wyższe u chorych z podtypem siateczkowatym niż u zdrowych osób z grupy kontrolnej [30]. Obserwacja ta wskazuje na udział Th1-zależnych reakcji zapalnych w procesie patogenezy.
Transformujący czynnik wzrostu β (TGF-β)
Rola TGF-β w zakresie kancerogenezy jest dwojaka: we wczesnych stadiach może hamować wzrost guza, natomiast w stadiach bardziej zaawansowanych może sprzyjać rozwojowi nowotworu poprzez indukcję angiogenezy i aktywację MMP-9 [4]. Na podstawie tych ustaleń oraz zgodnie ze stanowiskiem WHO, że liszaj płaski jest schorzeniem o potencjalnie złośliwym charakterze, przeprowadzono kilka badań, których celem była analiza ekspresji TGF-β. Stwierdzono znamiennie obniżone stężenie TGF-β w surowicy [39]. Ocena immunohistochemiczna TGF-β1 wykazała także umiarkowanie dodatnią ekspresję w nabłonku chorych z liszajem płaskim [42]. Obserwowano ponadto znamiennie zwiększoną ekspresję TGF-β u chorych z liszajem płaskim w porównaniu z prawidłową błoną śluzową jamy ustnej oraz dodatnią zależność pomiędzy ekspresją TGF-β1 a apoptozą komórek nabłonka [42]. Wyniki te wskazują na istotną rolę TGF-β w patogenezie liszaja płaskiego i rozwoju SCC na podłożu zmian liszaja płaskiego.
Inne markery
MMP 2, 9, 4, 7
Metaloproteinazy macierzy pozakomórkowej (MMPs) powodują uszkodzenia błony podstawnej i apoptozę keratynocytów [7]. Wysoki poziom ekspresji MMPs stwierdza się u pacjentów z liszajem płaskim [7]. Rubaci i wsp. wykazali podwyższoną ekspresję MMP-2 i MMP-7 w nabłonku i tkance łącznej u pacjentów z liszajem płaskim w porównaniu z grupą kontrolną [4]. Inni badacze obserwowali znamienne różnice w stężeniu MMPs pomiędzy postacią nadżerkową a nienadżerkową. Podwyższone stężenia MMP-1, MMP-2, MMP-3 i MMP-4 były obecne w podtypie nadżerkowym liszaja płaskiego [4]. W badaniu Wang i wsp. miejscowa ekspresja MMP-9 była podwyższona u pacjentów z liszajem płaskim w porównaniu z grupą kontrolną [13]. Inni badacze stwierdzili znamiennie podwyższone stężenie MMP-9 w tkankach przyzębia w tej grupie pacjentów [13].
Ze względu na destrukcyjne właściwości MMP badano zależność między ich ekspresją w zmianach chorobowych a procesem transformacji złośliwej. Na podstawie uzyskanych wyników ustalono, że MMP-2 i MMP-9 stanowią potencjalne markery złośliwej transformacji liszaja płaskiego [4, 7].
MHC
W zmianach wywoływanych przez liszaj płaski stwierdzano nadekspresję MHC klasy I i II. Ponadto obserwowano, że blokowanie wyłącznie HLA-DR, w przeciwieństwie do HLA-DP i DQ, chroni przed śmiercią keratynocytów [43]. W przypadku blokowania MHC klasy I odnotowywano bardziej nasilone oddziaływanie hamujące na śmierć keratynocytów indukowaną przez limfocyty T, co wskazuje na udział MHC klasy I w prezentacji antygenu w liszaju płaskim [43].
Cząsteczka adhezji komórkowej będąca adresyną błon śluzowych 1 (MAdCAM-1)
Stężenie MAdCAM-1 było w sposób znamienny obniżone w subpopulacji pacjentów z liszajem płaskim. Stwierdzono, że MAdCAM-1 może mieć znaczenie jako wskaźnik stanu zapalnego, ale nie jako marker aktywności choroby [44].
Wnioski
W liszaju płaskim nie ustalono dotąd czynnika patogenetycznego odpowiadającego za rozwój tej choroby, jednak patogeneza schorzenia jest ściśle związana z zaburzeniami immunologicznymi. Nieprawidłowości immunologiczne obejmują zróżnicowaną populację komórek odpornościowych, co może wskazywać na istnienie wspólnej cząsteczki inicjującej ten proces. Interesująca jest obserwacja, że liczba komórek prezentujących antygen wzrasta w stadiach poprzedzających rozwój pełnoobjawowej choroby.
Znajomość odchyleń w stężeniach markerów immunologicznych, m.in. cytokiny w ślinie i surowicy, może być przydatna w precyzyjnej diagnostyce bez użycia metod inwazyjnych, takich jak biopsja. W postaci siateczkowatej liszaja płaskiego obserwowane jest podwyższone stężenie IL-25, IL-10 i IFN-g, natomiast w postaci nadżerkowej występuje zwiększone stężenie IL-4, IL-6, IL-6, IL-17, IL-22 i TNF. Analiza stężenia IL-6 w ślinie może być przydatna w ocenie stopnia zaawansowania choroby i monitorowaniu odpowiedzi na leczenie. Podwyższone stężenie IL-22 w surowicy może służyć jako marker nawrotu choroby. W przypadku remisji obserwowane są obniżone stężenia IL-17, IL-21 i IL-22.
Należy zawsze pamiętać, że z powodu przewlekłego stanu zapalnego liszaj płaski jest stanem przednowotworowym. Wyniki, takie jak podwyższone stężenie MMP-2 i MMP-9 oraz zmniejszone stężenie TGF-β, sygnalizują wzrost potencjalnego ryzyka transformacji nowotworowej.
Przedstawione powyżej ustalenia mogą być przydatne w opracowaniu skutecznego leczenia przyczynowego, szczególnie w przypadkach opornych na standardową terapię. Diagnostyka liszaja płaskiego na podstawie markerów immunologicznych jest nowym obszarem dermatologii. Ze względu na ograniczoną dotąd liczbę badań w tym zakresie nie zaleca się stosowania tej metody jako jedynego narzędzia diagnostycznego. Potrzebne są dalsze badania i obserwacje, a pomimo obiecujących wyników nie można na razie wyeliminować złotego standardu diagnostycznego – badania histopatologicznego.
Konflikt interesów
Autorki nie zgłaszają konfliktu interesów.
Piśmiennictwo
1. Usatine R.P., Tinitigan M.: Diagnosis and treatment of lichen planus. Am Fam Physician 2011, 84, 53-60.
2. Shan J., Shen C., Fang J., Li S., Fan Y.: Potential roles of the CCL17-CCR4 axis in the immunopathogenesis of oral lichen planus. J Oral Pathol Med 2020, 49, 328-334.
3. Mozaffari H.R., Sharifi R., Hayati M., Imani M.M., Lopez-Jornet P., Golshah A., et al.: Evaluation of serum and salivary interferon-γ levels in patients with oral lichen planus: a systematic review and meta-analysis of case-control studies. Oral Surg Oral Med Oral Pathol Oral Radiol 2019, 127, 210-217.
4. Payeras M.R., Cherubini K., Figueiredo M.A., Salum F.G.: Oral lichen planus: focus on etiopathogenesis. Arch Oral Biol 2013, 58, 1057-1069.
5. Pekiner F.N., Demirel G.Y., Borahan M.O., Ozbayrak S.: Cytokine profiles in serum of patients with oral lichen planus. Cytokine 2012, 60, 701-706.
6. Humberto J.S.M., Pavanin J.V., Rocha M., Motta A.C.F.: Cytokines, cortisol, and nitric oxide as salivary biomarkers in oral lichen planus: a systematic review. Braz Oral Res 2018, 32, e82.
7. Mignogna M.D., Fedele S., Lo Russo L., Lo Muzio L., Bucci E.: Immune activation and chronic inflammation as the cause of malignancy in oral lichen planus: is there any evidence? Oral Oncol 2004, 40, 120-130.
8. Melguizo-Rodriguez L., Costela-Ruiz V.J., Manzano-Moreno F.J., Ruiz C., Illescas-Montes R.: Salivary biomarkers and their application in the diagnosis and monitoring of the most common oral pathologies. Int J Mol Sci 2020, 21, 5173.
9. Carvalho M., Cavalieri D., Do Nascimento S., Lourenço T.G.B., Ramos D.V.R., Pasqualin D.D.C., et al.: Cytokines levels and salivary microbiome play a potential role in oral lichen planus diagnosis. Sci Rep 2019, 9, 18137.
10. Sinon S.H., Rich A.M., Firth N.A., Seymour G.J.: Does the toll-like receptor 2 (TLR2) play a role in oral mucosal lichen planus (OMLP)? Pathology 2011, 43, S65.
11. Mattila R., Ahlfors E., Syrjänen S.: CD27 and CD38 lymphocytes are detected in oral lichen planus lesions. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2011, 111, 211-217.
12. Devi M., Vijayalakshmi D., Dhivya K., Janane M.: Memory T cells (CD45RO) role and evaluation in pathogenesis of lichen planus and lichenoid mucositis. J Clin Diagn Res 2017, 11, ZC84-ZC6.
13. Wang H., Guan X., Luo Z., Liu Y., Ren Q., Zhao X.: The association and potentially destructive role of Th9/IL-9 is synergistic with Th17 cells by elevating MMP9 production in local lesions of oral lichen planus. J Oral Pathol Med 2018, 47, 425-433.
14. Payeras M.R., Cherubini K., Figueiredo M.A., Salum F.G.: Oral lichen planus: focus on etiopathogenesis. Arch Oral Biol 2013, 58, 1057-1069.
15. Enomoto A., Sato E., Yasuda T., Isomura T., Nagao T., Chikazu D.: Intraepithelial CD8+ lymphocytes as a predictive diagnostic biomarker for the remission of oral lichen planus. Human Pathol 2018, 74, 43-53.
16. Mignogna M.D., Fedele S., Lo Russo L., Lo Muzio L., Bucci E.: Immune activation and chronic inflammation as the cause of malignancy in oral lichen planus: is there any evidence ? Oral Oncol 2004, 40, 120-130.
17. Jablonska E., Garley M., Surazynski A., Grubczak K., Iwaniuk A., Borys J., et al.: Neutrophil extracellular traps (NETs) formation induced by TGF-beta in oral lichen planus – possible implications for the development of oral cancer. Immunobiology 2020, 225, 151901.
18. Ramalingam S., Malathi N., Thamizhchelvan H., Sangeetha N., Rajan S.T.: Role of mast cells in oral lichen planus and oral lichenoid reactions. Autoimm Dis 2018, 2018, 7936564.
19. Wang Y., Shang S., Sun Q., Chen J., Du G., Nie H., et al.: Increased infiltration of CD11 c(+)/CD123(+) dendritic cell subsets and upregulation of TLR/IFN-α signaling participate in pathogenesis of oral lichen planus. Oral Surg Oral Med Oral Pathol Oral Radiol 2018, 125, 459-467.e2.
20. Cardozo A.L., Moura-Castro C., Figueiredo M., Cuzzi T., Ramos-e-Silva M.: Oral lichen planus and dermal dendrocytes. Actas Dermosifiliogr 2009, 100, 46-52.
21. de Carvalho G.C., Hirata F.Y.A., Domingues R., Figueiredo C.A., Zaniboni M.C., Pereira N.V., et al.: Up-regulation of HMGB1 and TLR4 in skin lesions of lichen planus. Arch Dermatol Res 2018, 310, 523-528.
22. Zhang J., Tan Y.Q., Wei M.H., Ye X.J., Chen G.Y., Lu R., et al.: TLR4-induced B7-H1 on keratinocytes negatively regulates CD4(+) T cells and CD8(+) T cells responses in oral lichen planus. Exp Dermatol 2017, 26, 409-415.
23. Lin S.C., Sun A., Wu Y.C., Chiang C.P.: Presence of anti-basal cell antibodies in oral lichen planus. J Am Acad Dermatol 1992, 26, 943-947.
24. Bramanti T.E., Dekker N.P., Lozada-Nur F., Sauk J.J., Regezi J.A.: Heat shock (stress) proteins and γδ T lymphocytes in oral lichen planus. Oral Surg Oral Med Oral Pathol Oral Radiol Endodontol 1995, 80, 698-704.
25. Liu J., Shi Q., Yang S., Wang Q., Xu J., Guo B.: The relationship between levels of salivary and serum interleukin-6 and oral lichen planus: a systematic review and meta-analysis. J Am Dental Assoc 2017, 148, 743-749.e9.
26. Mehrbani S.P., Motahari P., Azar F.P., Ahari M.A.: Role of interleukin-4 in pathogenesis of oral lichen planus: a systematic review. Med Oral Patol Oral Cirugia Bucal 2020, 25, e410-e415.
27. Kurchenko A., Drannik G., Rehuretska R., duBuske L.: Assessment of select serum cytokines TNF-a and IL-22 in oral lichen planus patients. Ann Allergy Asthma Immunol 2018, 121, 5 Suppl, S50.
28. Wang H., Jiang Y., Wang H., Luo Z., Wang Y., Guan X.: IL-25 promotes Th2-type reactions and correlates with disease severity in the pathogenesis of oral lichen planus. Arch Oral Biol 2019, 98, 115-121.
29. Zychowska M., Batycka-Baran A., Baran W.: Increased serum level and high tissue immunoexpression of interleukin 17 in cutaneous lichen planus: a novel therapeutic target for recalcitrant cases? Dis Markers 2020, 2020, 6521274.
30. Mozaffari H.R., Zavattaro E., Saeedi M., Lopez-Jornet P., Sadeghi M., Safaei M., et al.: Serum and salivary interleukin-4 levels in patients with oral lichen planus: a systematic review and meta-analysis. Oral Surg Oral Med Oral Pathol Oral Radiol 2019, 128, 123-131.
31. Liu W.Z., He M.J., Long L., Mu D.L., Xu M.S., Xing X., et al.: Interferon-γ and interleukin-4 detected in serum and saliva from patients with oral lichen planus. Int J Oral Sci 2014, 6, 22-26.
32. Dan H., Liu W., Wang J., Wang Z., Wu R., Chen Q., et al.: Elevated IL-10 concentrations in serum and saliva from patients with oral lichen planus. Quintessence Int 2011, 42, 157-163.
33. Negi D., Urs A.B., Kumar P., Mahajan B., Singh H., Polipalli S.K., et al.: Assessment of interleukin-18 gene polymorphism and serum levels in oral lichen planus in an Indian population. J Oral Pathol Med 2019, 48, 244-250.
34. Zhang Y., Liu W., Zhang S., Dan H., Lu R., Wang F., et al.: Salivary and serum interleukin-18 in patients with oral lichen planus: a study in an ethnic Chinese population. Inflammation 2012, 35, 399-404.
35. Drannik G.N., Kurchenko A.I., Rehuretska R.A., Drannik A.G., DuBuske L.M: Th-17 cytokines in oral lichen planus. J Allergy Clin Immunol 2014, 133, AB186.
36. Marshall A., Celentano A., Cirillo N., McCullough M., Porter S.: Tissue-specific regulation of CXCL9/10/11 chemokines in keratinocytes: implications for oral inflammatory disease. PLoS One 2017, 12, e0172821.
37. Fang J., Wang C., Shen C., Shan J., Wang X., Liu L., et al.: The expression of CXCL10/CXCR3 and effect of the axis on the function of T lymphocyte involved in oral lichen planus. Inflammation 2019, 42, 799-810.
38. Shan J., Li S., Wang C., Liu L., Wang X., Zhu D., et al.: Expression and biological functions of the CCL5-CCR5 axis in oral lichen planus. Exp Dermatol 2019, 28, 816-821.
39. Taghavi Zenouz A., Pouralibaba F., Babaloo Z., Mehdipour M., Jamali Z.: Evaluation of serum TNF-α and TGF-β in patients with oral lichen planus. J Dent Res Dent Clin Dent Prospects 2012, 6, 143-147.
40. Ding M., Zeng J., Sroussi H., Yu J., Xu J., Cheng X., et al.: Interactions between Golli-MBP and Th1/Th2 cytokines in patients with oral lichen planus. Oral Dis 2014, 20, 205-211.
41. Xie S., Ding L., Xiong Z., Zhu S.: Implications of Th1 and Th17 cells in pathogenesis of oral lichen planus. J Huazhong Univ Sci Technolog Med Sci 2012, 32, 451-457.
42. Chen Z.L., Deng G.H., Chen H.B., Cheng J.: Expression of TGF-beta1, Smad7 and cell apoptosis in epithelium of oral lichen planus. Zhonghua Kou Qiang Yi Xue Za Zhi 2008, 43, 95-98.
43. Shao S., Tsoi L.C., Sarkar M.K., Xing X., Xue K., Uppala R., et al.: IFN-γ enhances cell-mediated cytotoxicity against keratinocytes via JAK2/STAT1 in lichen planus. Sci Transl Med 2019, 11, eaav7561.
44. Baran A., Nowowiejska J., Kamiński T.W., Krahel J.A., Flisiak I.: Circulating MAdCAM-1 and ITGB7 in patients with plaque psoriasis and eruptive lichen planus-preliminary data. Biology 2021, 10, 1129.